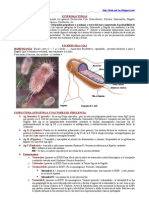
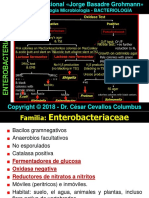
También podría gustarte
- Escherichia Coli, Salmonella, ShigellaDocumento14 páginasEscherichia Coli, Salmonella, ShigellacarmenAún no hay calificaciones
- Enterobacteriaceae CapDocumento11 páginasEnterobacteriaceae CapMaria GalgueraAún no hay calificaciones
- CoprocultivoDocumento5 páginasCoprocultivoManuel GilAún no hay calificaciones
- Micro Resumen 1 Unidad IiDocumento10 páginasMicro Resumen 1 Unidad IiAngie Pamela Mirella Colop PuacAún no hay calificaciones
- EnterobacteriasDocumento9 páginasEnterobacteriasAthina Borches MujicaAún no hay calificaciones
- Tema 8. Enterobacteriosis. Lección 15Documento8 páginasTema 8. Enterobacteriosis. Lección 15Ainoa Bersani RuizAún no hay calificaciones
- Trabajo BacteriologíaDocumento10 páginasTrabajo BacteriologíaWillinton Eraso BolañosAún no hay calificaciones
- Enterobacterias OportunistasDocumento26 páginasEnterobacterias OportunistasEstudiante de medicina0% (1)
- Procesamiento de CoprocultivosDocumento6 páginasProcesamiento de CoprocultivosQUIMICO CLINICO WILLIANS SANCHEZ100% (7)
- EnterobacteriasDocumento14 páginasEnterobacteriasPamela Ramos ParedesAún no hay calificaciones
- Cuestionario de Gram NegativasDocumento10 páginasCuestionario de Gram NegativasEdgar Josue Puicon BancesAún no hay calificaciones
- Enterobacterias, Marco Teórico. EmersonDocumento13 páginasEnterobacterias, Marco Teórico. EmersonEmerson NegronAún no hay calificaciones
- E. ColiDocumento17 páginasE. ColiAylinamvAún no hay calificaciones
- Escherichia ColiDocumento7 páginasEscherichia ColiWENDY PARRAGAAún no hay calificaciones
- E. Coli y Otras EnterobacteriasDocumento8 páginasE. Coli y Otras EnterobacteriasJosé Gabriel Quiroga VillagraAún no hay calificaciones
- Escherichia ColiDocumento6 páginasEscherichia ColiCarlos MirandaAún no hay calificaciones
- Caso Clinico InmunologiaDocumento7 páginasCaso Clinico Inmunologiafamilia cifuentes mendezAún no hay calificaciones
- UT 4-1 Bacteriologia de Los AlimentosDocumento56 páginasUT 4-1 Bacteriologia de Los AlimentosjosueAún no hay calificaciones
- E ColiDocumento7 páginasE ColiAdelisPuchurtintaIrcoAún no hay calificaciones
- Escherichia Coli EnterotoxigénicaDocumento17 páginasEscherichia Coli EnterotoxigénicaSabrina Reyna ValverdeAún no hay calificaciones
- P7 NMPcoliformes 19617Documento12 páginasP7 NMPcoliformes 19617Luis AguilarAún no hay calificaciones
- Escherichia ColiDocumento25 páginasEscherichia ColiMaria Del MarAún no hay calificaciones
- La Escherichia ColiDocumento6 páginasLa Escherichia ColiLaleska Salinas FallaAún no hay calificaciones
- Bacilos Gram Negativos No EsporulariosDocumento10 páginasBacilos Gram Negativos No EsporulariosJANNISPOZOAún no hay calificaciones
- EnterobacteriasDocumento13 páginasEnterobacteriasAbigail VCAún no hay calificaciones
- Familia EnterobacteriaceaeDocumento17 páginasFamilia EnterobacteriaceaePaulina BarretoAún no hay calificaciones
- Infecciones Del Tracto Digestivo Estudiantes PDFDocumento28 páginasInfecciones Del Tracto Digestivo Estudiantes PDFlordgimli77Aún no hay calificaciones
- Eda BacterianaDocumento8 páginasEda BacterianaDayita PeciosaAún no hay calificaciones
- BIO150 Clase 13-Gram Negativo 2012 VIPDocumento77 páginasBIO150 Clase 13-Gram Negativo 2012 VIPDaniela Navarro SepúlvedaAún no hay calificaciones
- EnterobacteriasDocumento13 páginasEnterobacteriasAbigail VCAún no hay calificaciones
- Resumen E. ColiDocumento3 páginasResumen E. ColiGeo CercadoAún no hay calificaciones
- Prof. Jorge Damián Foronda USP Chimbote-2014Documento60 páginasProf. Jorge Damián Foronda USP Chimbote-2014KassandraAltamiranoAún no hay calificaciones
- Escherichia Spp.Documento24 páginasEscherichia Spp.Ninfa MoralesAún no hay calificaciones
- Bacterias 2Documento271 páginasBacterias 2Eglis KarolAún no hay calificaciones
- Bacilos GramnegativosDocumento12 páginasBacilos Gramnegativosangela milagros ramirez guerreroAún no hay calificaciones
- EDAS-GEPIB-SDA y CASOS CLIN Nov 25 de 2021Documento76 páginasEDAS-GEPIB-SDA y CASOS CLIN Nov 25 de 2021Mildred Andrea Olguin GonzalezAún no hay calificaciones
- Introduccion Tecnica NMP Coliformes Totales y FecalesDocumento3 páginasIntroduccion Tecnica NMP Coliformes Totales y FecalesMiguel Eduardo Arrieta MilanésAún no hay calificaciones
- wuolah-free-CUADRO TEMA 8 (Parte 2) PDFDocumento9 páginaswuolah-free-CUADRO TEMA 8 (Parte 2) PDFShay SinmasAún no hay calificaciones
- EnterobacteriaceaeDocumento91 páginasEnterobacteriaceaes7vdvm28btAún no hay calificaciones
- EnterobacteriasDocumento49 páginasEnterobacteriasAlejo Torres100% (1)
- Bacilos Gramnegativos. EnterobacteriasDocumento13 páginasBacilos Gramnegativos. EnterobacteriasAndreaFerreiraAún no hay calificaciones
- E ColiDocumento12 páginasE ColiCristian Tarazona Vargas100% (1)
- Tarea 2 de Micro 2 FaseDocumento18 páginasTarea 2 de Micro 2 FaseEduardo Oscar Quicaño YanaAún no hay calificaciones
- MICROBIOLOGIADocumento7 páginasMICROBIOLOGIAPeqe BustosAún no hay calificaciones
- E. Coli y SerotiposDocumento7 páginasE. Coli y SerotiposCarolinaJaimesAún no hay calificaciones
- EscherichiaDocumento4 páginasEscherichiaFernanda ChavezAún no hay calificaciones
- Analisis Microbiologico Del Agua en La Comunidad Del Poblado y Ejido El Veladero MpioDocumento14 páginasAnalisis Microbiologico Del Agua en La Comunidad Del Poblado y Ejido El Veladero MpioJoel Vlelez JuarezAún no hay calificaciones
- Escherichia ColiDocumento13 páginasEscherichia ColiJacky Tasayco100% (2)
- Bacilos Gram NegativosDocumento63 páginasBacilos Gram NegativosPaola Yanara VargasAún no hay calificaciones
- Bacterias PatogenasDocumento34 páginasBacterias PatogenasDiegoAún no hay calificaciones
- EnterobacteriasDocumento52 páginasEnterobacteriasPeter RGAún no hay calificaciones
- La Bacteria e ColiDocumento4 páginasLa Bacteria e ColiDavid Espinosa SuárezAún no hay calificaciones
- Infecciones Gastrointestinales 2023Documento50 páginasInfecciones Gastrointestinales 2023mateoAún no hay calificaciones
- Enterobacterias Medicine2010Documento6 páginasEnterobacterias Medicine2010Wiiny LissiAún no hay calificaciones
- Bacterias Gram NegativasDocumento7 páginasBacterias Gram NegativasPaola CadenaAún no hay calificaciones
- Patogenicidad de Escherichia ColiDocumento9 páginasPatogenicidad de Escherichia ColiLeonardoEscalanteAún no hay calificaciones
- ENTEROBACTERIASDocumento72 páginasENTEROBACTERIASLucasAún no hay calificaciones
- EnterobacteriasDocumento10 páginasEnterobacteriasbilbao.pilar1Aún no hay calificaciones
- Los bacteriófagos: Los virus que combaten infeccionesDe EverandLos bacteriófagos: Los virus que combaten infeccionesAún no hay calificaciones
- Diarrea Infecciosa AgudaDocumento118 páginasDiarrea Infecciosa AgudaPaolaAlejandraPaucarRomeroAún no hay calificaciones
- Guía 1 Tecnologia e Inform 9Documento4 páginasGuía 1 Tecnologia e Inform 9Yeribeth LanderosAún no hay calificaciones
- 9 YersiniaDocumento6 páginas9 YersiniaAntonio RomanAún no hay calificaciones
- Bacilos Gram NegativosDocumento34 páginasBacilos Gram NegativosAntonyAún no hay calificaciones
- Yersinia EnterocoliticaDocumento10 páginasYersinia EnterocoliticaCarolineReyesAlcántara0% (1)
- Resumen de YersiniaDocumento2 páginasResumen de YersiniaJosué CandelarioAún no hay calificaciones
- 272 Infecciones Por Salmonella y Yersinia PDFDocumento11 páginas272 Infecciones Por Salmonella y Yersinia PDFliancaAún no hay calificaciones
- Bacilos Gram NegativosDocumento63 páginasBacilos Gram NegativosgabypintoAún no hay calificaciones
- Paradigma de La Microbiologia HistoriaDocumento8 páginasParadigma de La Microbiologia HistoriaAlejandraAún no hay calificaciones
- Cuadro de Doble EntradaDocumento3 páginasCuadro de Doble Entradadiana lisbeth bustamante valdiviaAún no hay calificaciones
- Yersinia EnterocolíticaDocumento2 páginasYersinia EnterocolíticaFabricio Melgarejo AlvaradoAún no hay calificaciones
- BacteriasDocumento8 páginasBacteriasFIDALGO JOSE LOPEZ VELASQUEZAún no hay calificaciones
- TEMA - 9 - PARTE - IIc Bacilos GramDocumento39 páginasTEMA - 9 - PARTE - IIc Bacilos GramUYIAún no hay calificaciones
- Yersinia ExpoDocumento15 páginasYersinia ExpoEmy MerinoAún no hay calificaciones
- Resumen BacteriasDocumento15 páginasResumen BacteriasDaniel Alejandro Roa TovarAún no hay calificaciones
- Bacterias 2Documento9 páginasBacterias 2Perla CastilloAún no hay calificaciones
- Bacterias Que Afectan El Sistema DigestivoDocumento26 páginasBacterias Que Afectan El Sistema Digestivopixmar0% (3)
- Esquemas Microbiologia PDFDocumento8 páginasEsquemas Microbiologia PDFXanecouboi XikiAún no hay calificaciones
- Practica # 8Documento10 páginasPractica # 8MaryDakota KnightAún no hay calificaciones
- Yersenia y PasteurellaDocumento38 páginasYersenia y PasteurellaGisse May SocasiAún no hay calificaciones
- Resumen Bactérias - VagnerDocumento93 páginasResumen Bactérias - VagnerMayra LandoAún no hay calificaciones
- Clase # 9 AEROMONAS HYDROPHILADocumento10 páginasClase # 9 AEROMONAS HYDROPHILATamara VeraAún no hay calificaciones
- Yersinia PestisDocumento4 páginasYersinia PestispamelamalizaAún no hay calificaciones
- Proceso de Salud en Bolivia PSBDocumento12 páginasProceso de Salud en Bolivia PSBWaldo Jhasmani Lopez CruzAún no hay calificaciones
- Familia EnterobacteriumDocumento64 páginasFamilia EnterobacteriumPaolo BarredaAún no hay calificaciones
- Contaminación de Los AlimentosDocumento47 páginasContaminación de Los AlimentosRobinson Enrique Villamonte RubioAún no hay calificaciones
- Secuencia Del Genoma de Yersinia Pestis KIM ResumenDocumento3 páginasSecuencia Del Genoma de Yersinia Pestis KIM ResumenvanessaAún no hay calificaciones
- Ficha Aplicativa 6toDocumento10 páginasFicha Aplicativa 6toEinstiano EmcAún no hay calificaciones
- Castillo Velasco Delfa Analisis Peste Negra.Documento8 páginasCastillo Velasco Delfa Analisis Peste Negra.Angelita Castillo VelascoAún no hay calificaciones
- Bacterias Ii P 2021Documento13 páginasBacterias Ii P 2021Obed GodenAún no hay calificaciones