También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Acción de Los Agentes Humorales Sobre La Membrana y Sobre Los Órganos IntracitoplasmáticosDocumento14 páginasAcción de Los Agentes Humorales Sobre La Membrana y Sobre Los Órganos IntracitoplasmáticosFernando A. Gurrola50% (2)
- CLASIFICACIONPROGESTINASDocumento20 páginasCLASIFICACIONPROGESTINASEdwin PeraltaAún no hay calificaciones
- Receptor de InsulinaDocumento20 páginasReceptor de InsulinaIgnacioNacho Reyes0% (1)
- RECEPTORESDocumento5 páginasRECEPTORESMaryen Gallardo ovandoAún no hay calificaciones
- GlucoproteínasDocumento9 páginasGlucoproteínasUn pedacito de CieloAún no hay calificaciones
- Operon LactosaDocumento52 páginasOperon LactosaAmbar GabrielaAún no hay calificaciones
- Resumen GastrulacionDocumento2 páginasResumen Gastrulacionfireball764100% (2)
- 3 Adrenalina, Noradrenalina. 3ra ClaseDocumento88 páginas3 Adrenalina, Noradrenalina. 3ra ClaseRosa PaezAún no hay calificaciones
- AnovulacionDocumento18 páginasAnovulacionMailyn Guerrero PardoAún no hay calificaciones
- Regulación de La Actividad Enzimatica Grupo 3Documento29 páginasRegulación de La Actividad Enzimatica Grupo 3AlexandraParedesPuertaAún no hay calificaciones
- Ross. Cap 21Documento8 páginasRoss. Cap 21Patricia MartinsAún no hay calificaciones
- Hormonas y Señalización IntracelularDocumento49 páginasHormonas y Señalización IntracelularValmore Jose Bermudez100% (1)
- Mola Hidatiforme. Seminario 6 FisiologíaDocumento42 páginasMola Hidatiforme. Seminario 6 FisiologíaJosé Roberto Sánchez Méndez100% (1)
- GLUCOGENESISDocumento15 páginasGLUCOGENESISCamila Vargas VeraAún no hay calificaciones
- Adaptación Del Sistema HEPATICODocumento42 páginasAdaptación Del Sistema HEPATICOMaría Angelica DAppollonioAún no hay calificaciones
- Señalización Celular y Transducción de Señales FinalDocumento7 páginasSeñalización Celular y Transducción de Señales FinalLeón LopezAún no hay calificaciones
- Perfil GinecologicoDocumento18 páginasPerfil GinecologicoluzveliaAún no hay calificaciones
- La Secreción de La InsulinaDocumento5 páginasLa Secreción de La Insulinaeloydc17Aún no hay calificaciones
- Capítulo 15 - Señalización Celular y Transducción de Señales - Comunicación IntercelularDocumento75 páginasCapítulo 15 - Señalización Celular y Transducción de Señales - Comunicación IntercelularALEJANDRA INÉS NIETO ARIASAún no hay calificaciones
- Tema 6. LA MADURACIÓN DE LAS PROTEÍNASDocumento5 páginasTema 6. LA MADURACIÓN DE LAS PROTEÍNASNataly Garcia Navas0% (1)
- Interacción Fármaco Receptor y FarmacodinamiaDocumento27 páginasInteracción Fármaco Receptor y FarmacodinamiaJaqueline Gutierrez100% (1)
- RECEPTORES ACOPLADOS A LA PROTEINA G Practica IDocumento5 páginasRECEPTORES ACOPLADOS A LA PROTEINA G Practica IJose CarlosAún no hay calificaciones
- CRHDocumento3 páginasCRHMiguel Coa TitoAún no hay calificaciones
- Mecanismos de Regulación EnzimáticaDocumento2 páginasMecanismos de Regulación EnzimáticaRobert Isaias Mendoza CardenasAún no hay calificaciones
- Cadena Transportadora de Electrones y Fosforilacion OxidativaDocumento29 páginasCadena Transportadora de Electrones y Fosforilacion OxidativaLoreto Carrasco AlarconAún no hay calificaciones
- Señalización Celular - Matriz ExtracelularDocumento7 páginasSeñalización Celular - Matriz ExtracelularEnzo Guillermo RamirezAún no hay calificaciones
- Determinación de HormonasDocumento7 páginasDeterminación de HormonasSebastian RodasAún no hay calificaciones
- Anovulacion CronicaDocumento21 páginasAnovulacion CronicaTanDelValleAún no hay calificaciones
- Gestacion, Fecundacion, ImplantacionDocumento9 páginasGestacion, Fecundacion, ImplantacionFranklin TevesAún no hay calificaciones
- Sangre y HemostasiaDocumento57 páginasSangre y HemostasiaMacarena Riffo WestermannAún no hay calificaciones
- Antileucotrienos Mecanismo de AcciónDocumento2 páginasAntileucotrienos Mecanismo de AcciónAngelita Sarango100% (1)
- Enzimas Cardiacas CK CKMB Tgoasp LDHDocumento23 páginasEnzimas Cardiacas CK CKMB Tgoasp LDHAlberto GranadosAún no hay calificaciones
- EspermiogenesisDocumento3 páginasEspermiogenesisMariaJoseRenteriaCabrejos100% (3)
- Analogos GNRH CorrDocumento12 páginasAnalogos GNRH CorrCarolina RamosAún no hay calificaciones
- ESTEROIDOGENESIS YanniDocumento47 páginasESTEROIDOGENESIS YanniYannika RosalesAún no hay calificaciones
- División CelularDocumento10 páginasDivisión CelularAndrea Fernanda Calderon CuadrosAún no hay calificaciones
- Insulina GlucagonDocumento24 páginasInsulina GlucagonAugusto Polo Casale Lara0% (1)
- Compuestos NitrogenadosDocumento192 páginasCompuestos NitrogenadosMilton IsraelAún no hay calificaciones
- Proteina GDocumento6 páginasProteina GAngela Gonzales HerreraAún no hay calificaciones
- Receptores HormonalesDocumento9 páginasReceptores HormonalesEdgar Lopez Jimenez100% (1)
- Ciclo Ovárico y Ciclo MenstrualDocumento4 páginasCiclo Ovárico y Ciclo MenstrualDenisse SarabiaAún no hay calificaciones
- Metabolismo EritrocitoDocumento5 páginasMetabolismo Eritrocitofrankesm2011Aún no hay calificaciones
- Maduración PulmonarDocumento30 páginasMaduración PulmonarJonny CardenasAún no hay calificaciones
- Resumen Guyton y Hall CAPITULO 81Documento11 páginasResumen Guyton y Hall CAPITULO 81Daniel Jaramillo AponteAún no hay calificaciones
- Capítulo 62 GuytonDocumento31 páginasCapítulo 62 Guytonaike7Aún no hay calificaciones
- Caso Clínico FeocromocitomaDocumento35 páginasCaso Clínico FeocromocitomaVero CabreraAún no hay calificaciones
- Conjugación BacterianaDocumento2 páginasConjugación BacterianaLuis Andres Henriquez Pasten100% (1)
- El GloméruloDocumento15 páginasEl GloméruloHarumi Yonashiro SAún no hay calificaciones
- Fosforilación OxidativaDocumento4 páginasFosforilación OxidativaDaleska C. GalarzaAún no hay calificaciones
- Estrógenos y ProgestágenosDocumento24 páginasEstrógenos y ProgestágenosAriel Diaz SAún no hay calificaciones
- Seminario 4Documento10 páginasSeminario 4Paula100% (1)
- Anatomía de La YeguaDocumento15 páginasAnatomía de La YeguaPaula LealAún no hay calificaciones
- Toxicidad e Intoxicación Por FármacosDocumento29 páginasToxicidad e Intoxicación Por FármacosVanaii OrtiizAún no hay calificaciones
- Adrenocorticotropina o CorticotropinaDocumento4 páginasAdrenocorticotropina o CorticotropinaNora Cueva GuevaraAún no hay calificaciones
- Código GenéticoDocumento18 páginasCódigo Genéticopaz_saavedra_3Aún no hay calificaciones
- AnticoagulantesDocumento7 páginasAnticoagulantesLucas DuranAún no hay calificaciones
- Mecanismo de Acción de Los ReceptoresDocumento9 páginasMecanismo de Acción de Los ReceptoresPaola LaraAún no hay calificaciones
- 2 - Transducción de SeñalesDocumento5 páginas2 - Transducción de SeñalesHernán GuietAún no hay calificaciones
- Determinación de CreatininaDocumento5 páginasDeterminación de CreatininaJuan FuentesAún no hay calificaciones
- Ensayo de PancreasDocumento1 páginaEnsayo de PancreasJuan FuentesAún no hay calificaciones
- Manejo Del Paciente Psicótico DepresivoDocumento1 páginaManejo Del Paciente Psicótico DepresivoJuan FuentesAún no hay calificaciones
- Era Posgenómica en El Estudio de Los HelmintosDocumento3 páginasEra Posgenómica en El Estudio de Los HelmintosJuan FuentesAún no hay calificaciones
- Telangiectasia Hereditaria o Hemorragica Caracteristicas ClinicasDocumento4 páginasTelangiectasia Hereditaria o Hemorragica Caracteristicas ClinicasJuan FuentesAún no hay calificaciones
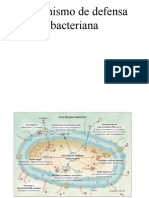
- Mecanismo de Defensa BacterianaDocumento3 páginasMecanismo de Defensa BacterianaJuan FuentesAún no hay calificaciones
- Abundancia Del OxigenoDocumento1 páginaAbundancia Del OxigenoJuan FuentesAún no hay calificaciones
- Preventiva Contra MosquitosDocumento2 páginasPreventiva Contra MosquitosJuan FuentesAún no hay calificaciones
- SentidosDocumento45 páginasSentidosJoos AlvaradoAún no hay calificaciones
- 3-Aminoacidos y ProteinasDocumento9 páginas3-Aminoacidos y ProteinasJuan FuentesAún no hay calificaciones
- Cuadernillo de Trabajo BiologiaDocumento28 páginasCuadernillo de Trabajo BiologianataliaAún no hay calificaciones
- Preguntas TP Educacion para La SaludDocumento3 páginasPreguntas TP Educacion para La Saludjennifer Comas100% (1)
- Tarea 6 - Raiven Smith AriasDocumento5 páginasTarea 6 - Raiven Smith AriasRuby EspinelAún no hay calificaciones
- Informe. FSCDocumento8 páginasInforme. FSCYajaira SeguraAún no hay calificaciones
- Fausto-Sterling - Cuerpos Sexuados-Páginas SeleccionadasDocumento9 páginasFausto-Sterling - Cuerpos Sexuados-Páginas SeleccionadasAnonymous 0eAfcg8PAún no hay calificaciones
- El ADN Portador Del Mensaje GenéticoDocumento20 páginasEl ADN Portador Del Mensaje GenéticoEsther MorrásAún no hay calificaciones
- Preguntas Biología Unidad 2Documento4 páginasPreguntas Biología Unidad 2HILDA BERNAL GONZALEZAún no hay calificaciones
- Informe - IDENTIFICACIÓN DE AMINOÁCIDOS Y PROTEÍNASDocumento12 páginasInforme - IDENTIFICACIÓN DE AMINOÁCIDOS Y PROTEÍNASalfredo andaluzAún no hay calificaciones
- MITOSIS UltimoDocumento15 páginasMITOSIS UltimojeisAún no hay calificaciones
- 2-Secuencia Modular IvDocumento30 páginas2-Secuencia Modular IvJorge Leonardo Ascencio CornelioAún no hay calificaciones
- Crucigrama Biologia ...Documento3 páginasCrucigrama Biologia ...leydy nataliaAún no hay calificaciones
- Actividad 7 Genetica y ComportamientoDocumento8 páginasActividad 7 Genetica y ComportamientoCARLOS ANDRES RAMIREZ CASTILLOAún no hay calificaciones
- Examen Simulado - Eci Área: BiologíaDocumento4 páginasExamen Simulado - Eci Área: BiologíaELBA ANGELA CUSICANQUI MALDONADOAún no hay calificaciones
- Cabeza Y CuelloDocumento36 páginasCabeza Y CuelloAdriana KummAún no hay calificaciones
- Consulta Previa 2Documento5 páginasConsulta Previa 2CRISTIAN DANIEL JIMÉNEZ CRUZ0% (1)
- PROTEINASDocumento11 páginasPROTEINASVirginia MorilloAún no hay calificaciones
- Dermoscromias Libro RepairedDocumento35 páginasDermoscromias Libro RepairedGénesis LeónAún no hay calificaciones
- Desarrollo Embrionario - Etapas y Sus Características (Semana A Semana) - LifederDocumento15 páginasDesarrollo Embrionario - Etapas y Sus Características (Semana A Semana) - LifederMevis Paola Ruiz JuncoAún no hay calificaciones
- 01 Bernard Stiegler, Lo - Inorganico - OrganizadoDocumento4 páginas01 Bernard Stiegler, Lo - Inorganico - OrganizadoDiego Fernando Arenas100% (1)
- Trabajo de Biología 3Documento1 páginaTrabajo de Biología 3Kevin PintagAún no hay calificaciones
- Expresion Genetica Del CoronavirusDocumento6 páginasExpresion Genetica Del Coronavirusanthony marc geerlings visagaAún no hay calificaciones
- Pruebas Reconocimiento de PaternidadDocumento4 páginasPruebas Reconocimiento de PaternidadFrancisco Barrón RochaAún no hay calificaciones
- Pep 1 Todas Las ClasesDocumento17 páginasPep 1 Todas Las ClasesJuan RPAún no hay calificaciones
- Bases Genéticas Del FitomejoramientoDocumento56 páginasBases Genéticas Del FitomejoramientoNayelhi carolina Valenzuela roncerosAún no hay calificaciones
- Gecko de Lima en Mateo Salado - Oportunidad Conservación PDFDocumento4 páginasGecko de Lima en Mateo Salado - Oportunidad Conservación PDFLauraPatriciaIglesiasCh.Aún no hay calificaciones
- Musculos Del Cuerpo HumanoDocumento8 páginasMusculos Del Cuerpo HumanoGENESIS CASTROAún no hay calificaciones
- Taller-Adaptacion (1) SisisiDocumento7 páginasTaller-Adaptacion (1) SisisiKamilita KamilitaAún no hay calificaciones
- Biologia SolomonDocumento3 páginasBiologia SolomonTerapia OcupacionalAún no hay calificaciones
- DesarrollóDocumento3 páginasDesarrollóOmarpimitty OmarpimittyAún no hay calificaciones
- 02 - MOLÉCULAS RECONOCEDORAS DE ANTÍGENOS y ANTÍGENOSDocumento4 páginas02 - MOLÉCULAS RECONOCEDORAS DE ANTÍGENOS y ANTÍGENOSLayla María Bebert BarrososAún no hay calificaciones
- Psicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaDe EverandPsicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaCalificación: 4.5 de 5 estrellas4.5/5 (766)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaDe EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaCalificación: 5 de 5 estrellas5/5 (201)
- Tus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadDe EverandTus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadCalificación: 4.5 de 5 estrellas4.5/5 (1831)
- Recupera tu mente, reconquista tu vidaDe EverandRecupera tu mente, reconquista tu vidaCalificación: 5 de 5 estrellas5/5 (6)
- Cómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaDe EverandCómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaCalificación: 5 de 5 estrellas5/5 (1870)
- Yo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)De EverandYo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)Calificación: 5 de 5 estrellas5/5 (7)
- Teoría polivagal práctica y terapiaDe EverandTeoría polivagal práctica y terapiaCalificación: 5 de 5 estrellas5/5 (4)
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- Resetea tu mente. Descubre de lo que eres capazDe EverandResetea tu mente. Descubre de lo que eres capazCalificación: 5 de 5 estrellas5/5 (196)
- El poder del optimismo: Herramientas para vivir de forma más positivaDe EverandEl poder del optimismo: Herramientas para vivir de forma más positivaCalificación: 5 de 5 estrellas5/5 (16)
- ¿Por qué mis padres no me aman?: Empezando a sanarDe Everand¿Por qué mis padres no me aman?: Empezando a sanarCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Influencia. La psicología de la persuasiónDe EverandInfluencia. La psicología de la persuasiónCalificación: 4.5 de 5 estrellas4.5/5 (14)
- Tu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaDe EverandTu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaCalificación: 5 de 5 estrellas5/5 (2)
- Una mente en calma: Técnicas para manejar los pensamientos intrusivosDe EverandUna mente en calma: Técnicas para manejar los pensamientos intrusivosCalificación: 4.5 de 5 estrellas4.5/5 (142)
- Terapia cognitivo-conductual (TCC) y terapia dialéctico-conductual (TDC): Cómo la TCC, la TDC y la ACT pueden ayudarle a superar la ansiedad, la depresión, y los TOCSDe EverandTerapia cognitivo-conductual (TCC) y terapia dialéctico-conductual (TDC): Cómo la TCC, la TDC y la ACT pueden ayudarle a superar la ansiedad, la depresión, y los TOCSCalificación: 5 de 5 estrellas5/5 (1)
- Cómo terminar lo que empiezas: El arte de perseverar, pasar a la acción, ejecutar los planes y tener disciplinaDe EverandCómo terminar lo que empiezas: El arte de perseverar, pasar a la acción, ejecutar los planes y tener disciplinaCalificación: 4.5 de 5 estrellas4.5/5 (6)
- Zensorialmente : Dejá que tu cuerpo sea tu cerebroDe EverandZensorialmente : Dejá que tu cuerpo sea tu cerebroAún no hay calificaciones
- No desperdicies tus emociones: Cómo lo que sientes te acerca a Dios y le da gloriaDe EverandNo desperdicies tus emociones: Cómo lo que sientes te acerca a Dios y le da gloriaCalificación: 5 de 5 estrellas5/5 (12)
- Disciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreDe EverandDisciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreCalificación: 5 de 5 estrellas5/5 (3)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (64)
- Las Cinco Leyes Biológicas De La Naturaleza: La Nueva Medicina (Color Edition) SpanishDe EverandLas Cinco Leyes Biológicas De La Naturaleza: La Nueva Medicina (Color Edition) SpanishCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Suicidología: Prevención, tratamiento psicológico e investigación de procesos suicidasDe EverandSuicidología: Prevención, tratamiento psicológico e investigación de procesos suicidasCalificación: 5 de 5 estrellas5/5 (7)
- Trucos para Leer la Mente de los Demás: Cómo Adivinar el Pensamiento de los Demás con Poco Esfuerzo. 2 Libros en 1 - Secretos de la Psicología Oscura, Cómo ser un Detector de MentirasDe EverandTrucos para Leer la Mente de los Demás: Cómo Adivinar el Pensamiento de los Demás con Poco Esfuerzo. 2 Libros en 1 - Secretos de la Psicología Oscura, Cómo ser un Detector de MentirasCalificación: 4.5 de 5 estrellas4.5/5 (3)