También podría gustarte
- Neuro Lenguaje y Ritmo CircardianoDocumento19 páginasNeuro Lenguaje y Ritmo CircardianoAlex LopezAún no hay calificaciones
- Neurobiología Del Sistema CircadianoDocumento11 páginasNeurobiología Del Sistema CircadianopsychforallAún no hay calificaciones
- Biorritmo y SexualidadDocumento19 páginasBiorritmo y SexualidadYass SiufiAún no hay calificaciones
- Circulación SistémicaDocumento7 páginasCirculación SistémicaCriisthii HernandezAún no hay calificaciones
- PNL TrabajoDocumento37 páginasPNL Trabajoibicelis33Aún no hay calificaciones
- Dhea Hormona JuventudDocumento7 páginasDhea Hormona JuventudBravo EbsAún no hay calificaciones
- Estudio Por Imagen Enfermedad CrohnDocumento14 páginasEstudio Por Imagen Enfermedad CrohnDiana Rioseras SardonAún no hay calificaciones
- Test de Imaginacion Con Base Al Amor InconscienteDocumento3 páginasTest de Imaginacion Con Base Al Amor InconscienteavunahAún no hay calificaciones
- Informe Histologia de La Cuerda VocalDocumento9 páginasInforme Histologia de La Cuerda VocalPaola Barria0% (1)
- Cronoterapia en PatologiasDocumento7 páginasCronoterapia en PatologiasBlanca Bayona ReinosoAún no hay calificaciones
- Glucosa PostprandialDocumento1 páginaGlucosa PostprandialAlbert Christopher Vargas QuirozAún no hay calificaciones
- Shock CardiogenicoDocumento47 páginasShock CardiogenicoTerapia IntensivaAún no hay calificaciones
- El Régimen Ancestral Del DRDocumento19 páginasEl Régimen Ancestral Del DRPedro FigueredoAún no hay calificaciones
- El Microbioma en Enfermedades AutoinmunesDocumento17 páginasEl Microbioma en Enfermedades AutoinmunesKevin OrtizAún no hay calificaciones
- Los Caminos de La CreaciónDocumento96 páginasLos Caminos de La CreaciónNicolás CaglieroAún no hay calificaciones
- Mecanismos de DetoxificaciónDocumento3 páginasMecanismos de DetoxificaciónEilyn CorralesAún no hay calificaciones
- Clase Miomatosis Uterina 2020Documento76 páginasClase Miomatosis Uterina 2020andy zavaleta100% (1)
- Músculo Esquelético Como Órgano EndocrinoDocumento43 páginasMúsculo Esquelético Como Órgano EndocrinoGabriela Carolina RamisAún no hay calificaciones
- Enfermedades Infecciosas y La Vit CDocumento8 páginasEnfermedades Infecciosas y La Vit CPablo Matas SoriaAún no hay calificaciones
- Biod UlanDocumento6 páginasBiod Ulancristal-liquido4191Aún no hay calificaciones
- PRONAP - Desnutrición GraveDocumento26 páginasPRONAP - Desnutrición GraveflorAún no hay calificaciones
- Plantas de La InsulinaDocumento17 páginasPlantas de La InsulinaSebastianAún no hay calificaciones
- AFMO 36 La Importancia Detoxificación C.A.Documento6 páginasAFMO 36 La Importancia Detoxificación C.A.Amparo Perez FerrerAún no hay calificaciones
- Endocrino UNIDAD VIDocumento31 páginasEndocrino UNIDAD VIAna Arcienega100% (1)
- MODULO IV. Glandula PinealDocumento11 páginasMODULO IV. Glandula PinealDarwinson RodriguezAún no hay calificaciones
- Vitaminas y AntioxidantesDocumento33 páginasVitaminas y AntioxidantesEfrain GonzalesAún no hay calificaciones
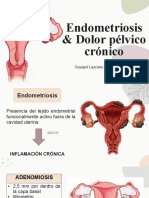
- Endometriosis y Dolor Pélvico CrónicoDocumento84 páginasEndometriosis y Dolor Pélvico CrónicoNaly Melissa Perez BastianAún no hay calificaciones
- Patologia de HigadoDocumento88 páginasPatologia de HigadojssscarlAún no hay calificaciones
- Enfermedad de Crohn (Autoguardado)Documento100 páginasEnfermedad de Crohn (Autoguardado)Jose Lopez FuentesAún no hay calificaciones
- Plantas Medicinales Del EcuadorDocumento35 páginasPlantas Medicinales Del Ecuadorjonatyhan maldonadoAún no hay calificaciones
- Ritmo CircadianoDocumento3 páginasRitmo CircadianoguaesAún no hay calificaciones
- Poliposdecolonyrecto 140515170116 Phpapp01 RemovedDocumento24 páginasPoliposdecolonyrecto 140515170116 Phpapp01 RemovedDouglas Bustamante100% (1)
- Apuntes de HematologiaDocumento65 páginasApuntes de HematologiaCitlaly MierAún no hay calificaciones
- Hormonas PDFDocumento27 páginasHormonas PDFMICHELLE VALDIVIA BIAGINIAún no hay calificaciones
- Genómica NutricionalDocumento18 páginasGenómica NutricionalCarlos RamírezAún no hay calificaciones
- Hildegarda de Bingen y Su Conocimiento CannabicoDocumento13 páginasHildegarda de Bingen y Su Conocimiento CannabicoSantiago CatólicoAún no hay calificaciones
- Como Alibiar El Dolor Menstrual NaturalmenteDocumento2 páginasComo Alibiar El Dolor Menstrual NaturalmenteFernando VegaAún no hay calificaciones
- 4 Clase Cancer de ProstataDocumento40 páginas4 Clase Cancer de ProstataSandra Zarate GonzalesAún no hay calificaciones
- Insuficiencia SuprarrenalDocumento3 páginasInsuficiencia SuprarrenalAna PaulaAún no hay calificaciones
- Consenso Nacional de Dieta Atkins ModificadaDocumento6 páginasConsenso Nacional de Dieta Atkins ModificadaPablo Antonio AntonioAún no hay calificaciones
- Exposición Cerebro EmocionalDocumento25 páginasExposición Cerebro EmocionalDana Paola Macal TasiasAún no hay calificaciones
- Monografía Final de Postgrado de Ayurveda 2013Documento15 páginasMonografía Final de Postgrado de Ayurveda 2013Cecilia Robles100% (1)
- Tecnicas y ProcedimientosDocumento60 páginasTecnicas y ProcedimientosRafael Sauceda Lopez100% (1)
- El Lactato y Su Papel en El Equilibrio Acido-BaseDocumento8 páginasEl Lactato y Su Papel en El Equilibrio Acido-Basevrllop100% (1)
- 5.apuntando Al Intestino para Tratar La Esclerosis MultipleDocumento21 páginas5.apuntando Al Intestino para Tratar La Esclerosis MultipleLourdes RuizAún no hay calificaciones
- Ascitis Diagnostico y ManejoDocumento34 páginasAscitis Diagnostico y ManejoJhon CastilloAún no hay calificaciones
- Comienzo y Evolución de La Enfermedad ActualDocumento9 páginasComienzo y Evolución de La Enfermedad ActualFlor FonsecaAún no hay calificaciones
- Cannabis y Cannabinoides Informacion para Profesionales de La Salud Canada 2013 en EspanolDocumento220 páginasCannabis y Cannabinoides Informacion para Profesionales de La Salud Canada 2013 en EspanolLorena PueblaAún no hay calificaciones
- RAPD Online 2009 V32 N2 07Documento14 páginasRAPD Online 2009 V32 N2 07Vicepresidenta ASOCEM UNLAún no hay calificaciones
- Afrodisiacos y AnafrodisiacosDocumento2 páginasAfrodisiacos y AnafrodisiacosMarian de Sánchez50% (2)
- Oximetria Pre y Post DuctualDocumento19 páginasOximetria Pre y Post DuctualDaissy Carolina Curmen DíazAún no hay calificaciones
- Electrofisiologia y Contraccion MuscularDocumento5 páginasElectrofisiologia y Contraccion Muscularjuan dominguezAún no hay calificaciones
- Dieta para El Reflujo GástricoDocumento6 páginasDieta para El Reflujo GástricoVania Marisella Mio RoqueAún no hay calificaciones
- Factores de Riesgo de Diabetes Mellitus Tipo 2 en Adultos de 18 A 59 Años en Hospital Sergio Bernales - Lima - Perú.Documento15 páginasFactores de Riesgo de Diabetes Mellitus Tipo 2 en Adultos de 18 A 59 Años en Hospital Sergio Bernales - Lima - Perú.Jesus Yampier Diaz ZeladaAún no hay calificaciones
- Atlas de Alimentos y Preparaciones 2018 PDFDocumento134 páginasAtlas de Alimentos y Preparaciones 2018 PDFAdrian ChurruarínAún no hay calificaciones
- UreaDocumento1 páginaUreaDavid Gómez AvalosAún no hay calificaciones
- Cronobiologia Funcion CardiovascularDocumento54 páginasCronobiologia Funcion CardiovascularJimmy Lui Che GallardoAún no hay calificaciones
- Ciclos CircadianosDocumento8 páginasCiclos Circadianosjuan david garces barrazaAún no hay calificaciones
- Cronobiologia EndocrinologiaDocumento16 páginasCronobiologia EndocrinologiacatatoumarocoAún no hay calificaciones
- Ritmo CircadianoDocumento5 páginasRitmo Circadianovladerdat100% (1)
- Mecanismos Circadianos en Medicina - NEJM 2021Documento22 páginasMecanismos Circadianos en Medicina - NEJM 2021EmanuelMCAún no hay calificaciones
- CTO 9ed - Ginecologia-ObstetriciaDocumento150 páginasCTO 9ed - Ginecologia-ObstetriciaEmanuelMC100% (1)
- No Todo Es ClínicaDocumento9 páginasNo Todo Es ClínicaEmanuelMCAún no hay calificaciones
- Internado Rotatorio. Medicina Interna 2017Documento768 páginasInternado Rotatorio. Medicina Interna 2017EmanuelMCAún no hay calificaciones
- Caso Clínico NocardiosisDocumento3 páginasCaso Clínico NocardiosisEmanuelMCAún no hay calificaciones
- Leishmaniasis (Incompleto)Documento17 páginasLeishmaniasis (Incompleto)EmanuelMC100% (2)
- Manual de Salud AmbientalDocumento56 páginasManual de Salud AmbientalEmanuelMCAún no hay calificaciones
- POSTER Leishmaniasis ECUADORDocumento3 páginasPOSTER Leishmaniasis ECUADOREmanuelMCAún no hay calificaciones
- Trauma ColumnaDocumento58 páginasTrauma ColumnaEmanuelMCAún no hay calificaciones
- Enfermedades de La Serie Roja: AnemiasDocumento16 páginasEnfermedades de La Serie Roja: AnemiasEmanuelMCAún no hay calificaciones
- Semiologia GastrointestinalDocumento23 páginasSemiologia GastrointestinalEmanuelMCAún no hay calificaciones
- TP y TPTDocumento3 páginasTP y TPTEmanuelMCAún no hay calificaciones
- Leishmaniasis - InformeDocumento14 páginasLeishmaniasis - InformeEmanuelMCAún no hay calificaciones
- Inmunologia - Cap#3 - Sistema de ComplementoDocumento23 páginasInmunologia - Cap#3 - Sistema de ComplementoEmanuelMC100% (1)
- Ensayo de Parasitologia, Trypanosoma CruziDocumento25 páginasEnsayo de Parasitologia, Trypanosoma CruziEmanuelMCAún no hay calificaciones
- Sindrome Nefrótico y NefríticoDocumento43 páginasSindrome Nefrótico y NefríticoEmanuelMCAún no hay calificaciones
- BACTERIAS GRAM NEGATIVAS Enterobacterias ExposicionDocumento56 páginasBACTERIAS GRAM NEGATIVAS Enterobacterias ExposicionEmanuelMC100% (2)
- 10 Comidas para Diabéticos Con Sus Recetas - Vida Birdman Blog - VidaBirdmanDocumento9 páginas10 Comidas para Diabéticos Con Sus Recetas - Vida Birdman Blog - VidaBirdmanPame SoledadAún no hay calificaciones
- Sesión 9 - Diabetes, Obesidad y Síndrome Metabólico.Documento73 páginasSesión 9 - Diabetes, Obesidad y Síndrome Metabólico.Flor Mendoza QuispeAún no hay calificaciones
- Caso ADocumento2 páginasCaso APAULA CORTESAún no hay calificaciones
- 1 - Fundamentos de Patología Diabética. PDFDocumento34 páginas1 - Fundamentos de Patología Diabética. PDFfelipe barrigaAún no hay calificaciones
- Biologia Parte Del Cuerpo HumanoDocumento3 páginasBiologia Parte Del Cuerpo Humanoeugenio hernandezAún no hay calificaciones
- Ejemplo ReviewDocumento12 páginasEjemplo ReviewEugenio Rangel LeónAún no hay calificaciones
- Taller 6 FarmacoDocumento7 páginasTaller 6 FarmacoBELEN HERREROS RICHARDAún no hay calificaciones
- Regulación de La Glucosa GlucógenoDocumento4 páginasRegulación de La Glucosa Glucógenorubipavia21Aún no hay calificaciones
- Diabetes. 24 Nov 2021Documento86 páginasDiabetes. 24 Nov 2021Richard LoewenherzAún no hay calificaciones
- EXPOSICIÓNDocumento6 páginasEXPOSICIÓNjuan diego osoriohenaoAún no hay calificaciones
- Practica N.2 - Bioquimica - Glucosa Capilar y Serica - 1ADocumento8 páginasPractica N.2 - Bioquimica - Glucosa Capilar y Serica - 1AKaren MooAún no hay calificaciones
- Tesis - Prevencion de La DiabetesDocumento39 páginasTesis - Prevencion de La DiabetesmakeupwithnanAún no hay calificaciones
- Mac Exposiciones HigienismoDocumento38 páginasMac Exposiciones HigienismoPamela GaravitoAún no hay calificaciones
- La Coordinación HormonalDocumento11 páginasLa Coordinación HormonalLeo FlorezAún no hay calificaciones
- Banco de Preguntas CURSO HUITRONDocumento151 páginasBanco de Preguntas CURSO HUITRONAaron Elvir100% (1)
- Programa Educativo de PacientesDocumento38 páginasPrograma Educativo de PacientesKM Javier100% (1)
- RecuperacionDocumento14 páginasRecuperacionsantiago23205Aún no hay calificaciones
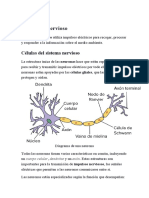
- El Sistema NerviosoDocumento7 páginasEl Sistema NerviosoMatias VillagraAún no hay calificaciones
- Mecanismos Bioquímicos de Los Receptores A Nivel Molecular JuditDocumento13 páginasMecanismos Bioquímicos de Los Receptores A Nivel Molecular Juditjesus neira100% (2)
- 18 InsulinasDocumento3 páginas18 Insulinasjen21morAún no hay calificaciones
- Entrevista - Rita - Charon PDFDocumento11 páginasEntrevista - Rita - Charon PDFAlejandra DiegoAún no hay calificaciones
- DEPICT-2 Study - Ensayo Clínico 2018Documento26 páginasDEPICT-2 Study - Ensayo Clínico 2018Freddy Shanner Chávez VásquezAún no hay calificaciones
- Diabetes MellitusDocumento73 páginasDiabetes MellitusFreddy Vargas RiveraAún no hay calificaciones
- Actividad Física Aeróbica y Anaeróbica.Documento7 páginasActividad Física Aeróbica y Anaeróbica.Jo-Nathan GrijalvaAún no hay calificaciones
- 25 - SHHDocumento11 páginas25 - SHHemmanuel antunezAún no hay calificaciones
- Fisiologia EndocrinaDocumento39 páginasFisiologia EndocrinaTomas Garciaguirre100% (1)
- Programa de SaludDocumento40 páginasPrograma de SaludChikitoOh Rivera SacramentoAún no hay calificaciones
- Pre Test - EndocrinologíaDocumento7 páginasPre Test - EndocrinologíaDaniel ChávezAún no hay calificaciones
- Guia Practicas Bioquimica Enfermeria Unt 2010Documento31 páginasGuia Practicas Bioquimica Enfermeria Unt 2010MariteYanceAún no hay calificaciones
- Monografia Diabetes Tipo IIDocumento32 páginasMonografia Diabetes Tipo IIMariano Trujillo ChávezAún no hay calificaciones