También podría gustarte
- Sistema EndocrinoDocumento88 páginasSistema EndocrinoLuis Fernando Rodríguez VillalpandoAún no hay calificaciones
- GUIA Sistema Endocrino II MedioDocumento7 páginasGUIA Sistema Endocrino II MedioREAL BUFONAún no hay calificaciones
- Hormonas Del Sistema EndocrinoDocumento10 páginasHormonas Del Sistema EndocrinoSteve Sanchez HAún no hay calificaciones
- EndocriinoDocumento5 páginasEndocriinoabelar216Aún no hay calificaciones
- Sistema Endocrino InformeDocumento8 páginasSistema Endocrino InformeFranciscoAún no hay calificaciones
- Tema 49. Introducción A La EndocrinologíaDocumento10 páginasTema 49. Introducción A La EndocrinologíaMarta AlonsoAún no hay calificaciones
- Sistemas Endocrino, Reproductor Masculino y FemeninoDocumento32 páginasSistemas Endocrino, Reproductor Masculino y FemeninoBrenda LopezAún no hay calificaciones
- HormonasDocumento10 páginasHormonasJaviera Escobedo CancinoAún no hay calificaciones
- UP 6 - FisiologiaDocumento27 páginasUP 6 - Fisiologiamaia càmporaAún no hay calificaciones
- Tutorial EndocrinoDocumento42 páginasTutorial Endocrinofrancis_bscAún no hay calificaciones
- Trabajo de Morfofisiologia II Cuido Mi Cuerpo Sistema EndocrinoDocumento24 páginasTrabajo de Morfofisiologia II Cuido Mi Cuerpo Sistema EndocrinoMary Hurtado MulethAún no hay calificaciones
- Sistema EndocrinoDocumento43 páginasSistema Endocrinoharmancek21Aún no hay calificaciones
- Clasificación de Las HormonasDocumento3 páginasClasificación de Las Hormonascarmen mogotocoro100% (1)
- Sistema EndocrinoDocumento5 páginasSistema EndocrinoPablo Daniel MontoyaAún no hay calificaciones
- P6. Hormonas. VereauDocumento31 páginasP6. Hormonas. VereauElmer MontenegroAún no hay calificaciones
- Sistema EndocrinoDocumento4 páginasSistema EndocrinoFabiola ZamudioAún no hay calificaciones
- Endocrino UagrmDocumento59 páginasEndocrino UagrmJean Carlos HerbasAún no hay calificaciones
- Marco Teórico EndocrinoDocumento9 páginasMarco Teórico EndocrinoPato Carmona Cortés50% (2)
- Hormonas IntroduccionDocumento27 páginasHormonas IntroduccionAnahi NavarroAún no hay calificaciones
- Introducción A La Endocrinología Fisio Cap 75Documento7 páginasIntroducción A La Endocrinología Fisio Cap 75liliana delgado67% (3)
- Caracteristicas HormonasDocumento9 páginasCaracteristicas HormonasFernando AntequeraAún no hay calificaciones
- Fisiología Del Sistema EndocrinoDocumento27 páginasFisiología Del Sistema EndocrinoAlba WAún no hay calificaciones
- FisioDocumento14 páginasFisioGabo GutierrezAún no hay calificaciones
- ENDOCRINOLOGIA 2019psicologiaDocumento6 páginasENDOCRINOLOGIA 2019psicologiaSofia GonzalezAún no hay calificaciones
- ENDOCRINOLOGÍADocumento14 páginasENDOCRINOLOGÍAharmancek21Aún no hay calificaciones
- Sistema-Endocrino 23Documento31 páginasSistema-Endocrino 23Ana Lorena Pérez MAún no hay calificaciones
- Guía Biología 2do C LUNES 04Documento5 páginasGuía Biología 2do C LUNES 04Paulina ZúñigaAún no hay calificaciones
- Sistema EndocrinoDocumento4 páginasSistema Endocrinoricardo matuteAún no hay calificaciones
- Organización Del Sistema Endócrino de Los MamíferosDocumento2 páginasOrganización Del Sistema Endócrino de Los MamíferosDIANAAún no hay calificaciones
- Teoría Del Sistema EndocrinoDocumento5 páginasTeoría Del Sistema EndocrinoFlor De Loto BlancoAún no hay calificaciones
- 1 Principios Generales de Fisiología EndocrinaDocumento17 páginas1 Principios Generales de Fisiología EndocrinaBarbara VegaAún no hay calificaciones
- Tipos de Hormonas y Sus Funciones en El Cuerpo HumanoDocumento13 páginasTipos de Hormonas y Sus Funciones en El Cuerpo HumanoPabloAún no hay calificaciones
- Tipos de Hormonas y Sus Funciones en El Cuerpo HumanoDocumento3 páginasTipos de Hormonas y Sus Funciones en El Cuerpo HumanoJosé Luis Ruiz100% (1)
- Sistema Endocrino 2023 UlDocumento5 páginasSistema Endocrino 2023 UlFranz Lazcano TarquiAún no hay calificaciones
- Clase 1 Fisio EndocrinaDocumento13 páginasClase 1 Fisio EndocrinaJeraldin SandovalAún no hay calificaciones
- 08 NeuroendocrinoDocumento30 páginas08 NeuroendocrinoAndrés Elías Londoño OquendoAún no hay calificaciones
- Guia Endocrino 2º Medio BioDocumento10 páginasGuia Endocrino 2º Medio Biokatherin rojasAún no hay calificaciones
- FISIOLOGI31Documento8 páginasFISIOLOGI31Maia MendozaAún no hay calificaciones
- Sistema Endocrino CCNN 2do.Documento25 páginasSistema Endocrino CCNN 2do.jose jesus Abraham gonzalez izaraAún no hay calificaciones
- Fisiologia Endocrina. Resumen y BancoDocumento62 páginasFisiologia Endocrina. Resumen y BancoZiulma Cano100% (5)
- Evaluación - MonohibridismoDocumento12 páginasEvaluación - MonohibridismoEvee VasquezAún no hay calificaciones
- Jesus Iglesia EndocrinoDocumento6 páginasJesus Iglesia Endocrinojenni de la peñaAún no hay calificaciones
- Sistema EndocrinoDocumento14 páginasSistema EndocrinoAnzoni Lara16Aún no hay calificaciones
- TALLER SISTEMA ENDOCRINO 2020 Convertido 2Documento15 páginasTALLER SISTEMA ENDOCRINO 2020 Convertido 2Yesenia Andrea Martinez HerreraAún no hay calificaciones
- Tarea Endocrino 2022Documento5 páginasTarea Endocrino 2022Madelin PinargoteAún no hay calificaciones
- EndocrinologíaDocumento42 páginasEndocrinologíajoaquín_navarro_6Aún no hay calificaciones
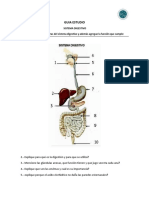
- Guía de EstudioDocumento5 páginasGuía de EstudioManuel Guerra PérezAún no hay calificaciones
- PsicoendocrinologiaDocumento5 páginasPsicoendocrinologiaYulet Melissa Aviles Lua0% (1)
- Generalidades Sistema EndocrinoDocumento30 páginasGeneralidades Sistema EndocrinoRaúl Cabrera100% (1)
- Tipos de HormonasDocumento2 páginasTipos de HormonasEmmanuel ValdezAún no hay calificaciones
- Wa0053.Documento41 páginasWa0053.Veronica OlivaresAún no hay calificaciones
- Universidad Autónoma de Durango: Campus Gomez Palacio DurangoDocumento14 páginasUniversidad Autónoma de Durango: Campus Gomez Palacio Durangosandy elizaldeAún no hay calificaciones
- Sistema EndocrinoDocumento46 páginasSistema EndocrinoMaria Jose Jara GuerreroAún no hay calificaciones
- Ross. Cap 21Documento8 páginasRoss. Cap 21Patricia MartinsAún no hay calificaciones
- Balance Hormonal/ Cómo Recuperar Balance Hormonal, Apetito Sexual, Sueño y Pérdida de Peso, AhoraDe EverandBalance Hormonal/ Cómo Recuperar Balance Hormonal, Apetito Sexual, Sueño y Pérdida de Peso, AhoraCalificación: 4 de 5 estrellas4/5 (1)
- Potenciadores De Testoterona: Descubre Los Suplementos Nutricionales Y Hierbas Utilizadas Para Mantener La Virilidad Toda La VidaDe EverandPotenciadores De Testoterona: Descubre Los Suplementos Nutricionales Y Hierbas Utilizadas Para Mantener La Virilidad Toda La VidaAún no hay calificaciones
- Aprenda Farmacología En Una SemanaDe EverandAprenda Farmacología En Una SemanaCalificación: 2 de 5 estrellas2/5 (1)
- Fisiologia de La Digestion en Los MonogastricosDocumento32 páginasFisiologia de La Digestion en Los MonogastricosDiana RAún no hay calificaciones
- Fisiologia V y VIDocumento29 páginasFisiologia V y VIDiana RAún no hay calificaciones
- Farmacología Veterinaria EduardoDocumento11 páginasFarmacología Veterinaria EduardoDiana RAún no hay calificaciones
- EndocrinologiaDocumento21 páginasEndocrinologiaDiana RAún no hay calificaciones
- Origen Del Lenguaje UNAM PDFDocumento7 páginasOrigen Del Lenguaje UNAM PDFIván BurroughsAún no hay calificaciones
- Verdadero Venezolano PDFDocumento51 páginasVerdadero Venezolano PDFJesús D. LeónAún no hay calificaciones
- Tarea1 - VanessaDaza MorfofisiologiaDocumento4 páginasTarea1 - VanessaDaza MorfofisiologiaJ&Jasesoriasacademicas AaAún no hay calificaciones
- ACTIVIDAD DE CLASE - SEMANA 1 (Anatomia y Fisiologia)Documento11 páginasACTIVIDAD DE CLASE - SEMANA 1 (Anatomia y Fisiologia)Claribel Sarait Blanco GiilAún no hay calificaciones
- Informe de Practica RIEGOS Y DRENAJESDocumento13 páginasInforme de Practica RIEGOS Y DRENAJESCEFACUNDOSAún no hay calificaciones
- ToxoplasmosisDocumento12 páginasToxoplasmosisAxel Trinidad LopezAún no hay calificaciones
- Algo Muy Importante Es Que El Niño .....Documento13 páginasAlgo Muy Importante Es Que El Niño .....Mery CaluchoAún no hay calificaciones
- 5to Año Plan de Estudio Area Biología I Momento Claudia T 2021-2022Documento4 páginas5to Año Plan de Estudio Area Biología I Momento Claudia T 2021-2022Gleydis SalazarAún no hay calificaciones
- Supernumeraria VertebraDocumento14 páginasSupernumeraria VertebraDiana Lorena Echeverri A100% (1)
- VIHDocumento16 páginasVIHbrayanAún no hay calificaciones
- Cinetica Inmovilizacion de EnzimasDocumento41 páginasCinetica Inmovilizacion de EnzimasJazmin PechAún no hay calificaciones
- LLANTENDocumento3 páginasLLANTENHeidi A FloresAún no hay calificaciones
- Oso Hormiguero El Regreso Del YurumiDocumento93 páginasOso Hormiguero El Regreso Del YurumiRoxmely GilAún no hay calificaciones
- Hidroterapia de ContrasteDocumento58 páginasHidroterapia de ContrasteJavier DiazAún no hay calificaciones
- Hemorragiasnn3n 5860da992c67f1fDocumento14 páginasHemorragiasnn3n 5860da992c67f1fMartha DonadoAún no hay calificaciones
- Meyer & Steinthal - Vocabulario Fundamental y Constructivo Del Griego PDFDocumento274 páginasMeyer & Steinthal - Vocabulario Fundamental y Constructivo Del Griego PDFGonzalo Diaz100% (1)
- Microsoft Word - Lombricultura Como Alternativa - Dr. Aguirre BortoniDocumento14 páginasMicrosoft Word - Lombricultura Como Alternativa - Dr. Aguirre BortoniCarol Vasquez HuamanAún no hay calificaciones
- Sonetos Del Crimen (Xorge Del Campo)Documento5 páginasSonetos Del Crimen (Xorge Del Campo)Alí Hassán FrancoAún no hay calificaciones
- El Primer PasoDocumento2 páginasEl Primer PasoDaniel CardenasAún no hay calificaciones
- Cuadernillo Biología - 3° ESODocumento149 páginasCuadernillo Biología - 3° ESOSandra Aguirre AngeloffAún no hay calificaciones
- Oms Criterios de Elegibilidad de Anticonceptivos 2015 FinalDocumento17 páginasOms Criterios de Elegibilidad de Anticonceptivos 2015 FinalFauryLlerenaAún no hay calificaciones
- 02 Prueba Batch Con Celda Neumatica Jameson - Marcelo AguilarDocumento22 páginas02 Prueba Batch Con Celda Neumatica Jameson - Marcelo AguilarRodrigo AlcainoAún no hay calificaciones
- Microbiota OralDocumento29 páginasMicrobiota OralSofia LainezAún no hay calificaciones
- Generalidades-de-Sistemática-AnimalDocumento12 páginasGeneralidades-de-Sistemática-AnimalIrving GomezAún no hay calificaciones
- CarbapenemsDocumento44 páginasCarbapenemsKevin Rodríguez Coronado100% (1)
- Las Ondas Alfa Del Electroencefalograma CuantificadoDocumento14 páginasLas Ondas Alfa Del Electroencefalograma CuantificadoAriel Rojas BarrientosAún no hay calificaciones
- Phylum RotiferaDocumento15 páginasPhylum RotiferaTatiana LaraAún no hay calificaciones
- Arbuzales y Herbazales Tepuyanos Resumen.Documento3 páginasArbuzales y Herbazales Tepuyanos Resumen.Yekarly MartinezAún no hay calificaciones
- Ciclo Biogeoquímico Del Nitrógeno Orientado A La SustentabilidadDocumento155 páginasCiclo Biogeoquímico Del Nitrógeno Orientado A La SustentabilidadAlberto Díaz0% (1)
- 2 Tejido EpitelialDocumento9 páginas2 Tejido EpitelialVeronica GonzalezAún no hay calificaciones