También podría gustarte
- Bases Biologicas de La Conducta CriminalDocumento13 páginasBases Biologicas de La Conducta CriminalJhonatan JalluranaAún no hay calificaciones
- Trastornos de La Función GlomerularDocumento20 páginasTrastornos de La Función GlomerularFabiola SánchezAún no hay calificaciones
- Qué Es Un GenDocumento1 páginaQué Es Un GenIrving Uriel Garcia CarranzaAún no hay calificaciones
- Encefalopatia Hepatica Lovesio PDFDocumento28 páginasEncefalopatia Hepatica Lovesio PDFFranklin López SalazarAún no hay calificaciones
- Crecimiento Poblacional de La LevaduraDocumento4 páginasCrecimiento Poblacional de La LevaduraJuan Jose MejiaAún no hay calificaciones
- Enzimas GeneralidadesDocumento28 páginasEnzimas GeneralidadesEdward SáenzAún no hay calificaciones
- Triptico AnticonceptivosDocumento2 páginasTriptico AnticonceptivosJosé Rafael50% (2)
- HemogramaDocumento29 páginasHemogramaDiego Iván De La Cruz100% (1)
- Darwin Ha Muerto... y Ahora QueDocumento288 páginasDarwin Ha Muerto... y Ahora QueTiempo InfinitoAún no hay calificaciones
- Triptico de Planificacion FamiliarDocumento2 páginasTriptico de Planificacion FamiliarJosé Rafael100% (4)
- FiebreDocumento1 páginaFiebreJosé Rafael100% (1)
- Gastroenteritis BacterianaDocumento30 páginasGastroenteritis BacterianaRuben SierraAún no hay calificaciones
- Ciclo Celular y CancerDocumento47 páginasCiclo Celular y CancerLesly Valencia CarrionAún no hay calificaciones
- FolletoDocumento2 páginasFolletoJosé RafaelAún no hay calificaciones
- Resumen PancreatitisDocumento14 páginasResumen PancreatitisKarina Itzel Castro EleuterioAún no hay calificaciones
- Diureticos-Isaza Ocr PDFDocumento10 páginasDiureticos-Isaza Ocr PDFSantiago Figueroa TorresAún no hay calificaciones
- ERITROCITOSDocumento38 páginasERITROCITOSCamila Andrea Villalobos AvilaAún no hay calificaciones
- Bioexpani Experimentacion Animal 2Documento209 páginasBioexpani Experimentacion Animal 2Francisco Jesus Querino SantiagoAún no hay calificaciones
- Introducción A La AntropologiaDocumento363 páginasIntroducción A La AntropologiaYasmira Barboza100% (6)
- Funcion HepaticaDocumento16 páginasFuncion HepaticaIgnacioAguilaBarradoAún no hay calificaciones
- 6th Central Pay Commission Salary CalculatorDocumento15 páginas6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- Bases Geneticas de Los Trastornos Del NeurodesarrolloDocumento12 páginasBases Geneticas de Los Trastornos Del NeurodesarrollororozainosAún no hay calificaciones
- LisosomasDocumento14 páginasLisosomasAndrea0% (1)
- Transporte de CalcioDocumento3 páginasTransporte de CalcioLuz FloresAún no hay calificaciones
- Fisiopatología-Insuficiencia HepaticaDocumento4 páginasFisiopatología-Insuficiencia HepaticaJuan Sebastian Montoya FernándezAún no hay calificaciones
- Nefrona DistalDocumento26 páginasNefrona DistalEddwwin Varillas Reyes100% (1)
- Trabajo RenalDocumento24 páginasTrabajo RenalBegglymar MedinaAún no hay calificaciones
- Sindrome de Mala AbsorcionDocumento3 páginasSindrome de Mala AbsorcionEbbi DvAún no hay calificaciones
- Reticulo Endoplasmático LisoDocumento4 páginasReticulo Endoplasmático LisoDel Valle CarlitosAún no hay calificaciones
- Diabetes InsipidaDocumento30 páginasDiabetes InsipidaEva KrEn SpAún no hay calificaciones
- La Nefrona, Tubulos Proximales y DistalesDocumento2 páginasLa Nefrona, Tubulos Proximales y DistalesHeidi Esther BaezAún no hay calificaciones
- GlosarioDocumento3 páginasGlosarioCeli ZarateAún no hay calificaciones
- Modelo de Atencion Integral de Salud Parte 2Documento3 páginasModelo de Atencion Integral de Salud Parte 2juan vasquezAún no hay calificaciones
- Tercer Parcial PATOLOGÍADocumento37 páginasTercer Parcial PATOLOGÍAVale Echavarri100% (1)
- Cuestionario de Glucidos y MetabolismoDocumento5 páginasCuestionario de Glucidos y MetabolismoIAGO SAMUEL DE SOUZA REISAún no hay calificaciones
- Pae 9Documento4 páginasPae 9ErîčkJavîêŕAún no hay calificaciones
- Alteraciones ElectroliticasDocumento52 páginasAlteraciones ElectroliticasJuanDavidMAún no hay calificaciones
- Patologia Del Sistema EndocrinoDocumento40 páginasPatologia Del Sistema EndocrinoRene Alexander Villalta VelasquezAún no hay calificaciones
- GLUCONEOGÉNESISDocumento10 páginasGLUCONEOGÉNESISNeydo PayaresAún no hay calificaciones
- Ruta de La Vida 4 DemorasDocumento44 páginasRuta de La Vida 4 DemorasMario Fernando Villota Esparza0% (1)
- Aparato de GolgiDocumento34 páginasAparato de GolgiMoisés J. Duran CanevaroAún no hay calificaciones
- Mecanismo de Concentración Por ContracorrienteDocumento2 páginasMecanismo de Concentración Por Contracorrientemi bella mas bellaAún no hay calificaciones
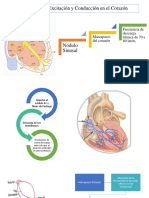
- Control de La Excitación y La Conducción Del CorazónDocumento10 páginasControl de La Excitación y La Conducción Del CorazónAnonymous VgRnIZEgtAún no hay calificaciones
- Tuberculosis RenalDocumento22 páginasTuberculosis Renalangel gutierrez100% (1)
- Cirrosis HepaticaDocumento12 páginasCirrosis HepaticaOscar Salvatierra diazAún no hay calificaciones
- Neumologia DiapositivasDocumento598 páginasNeumologia DiapositivasEdgar DoradoAún no hay calificaciones
- Covid Fisiopatologia - En.esDocumento21 páginasCovid Fisiopatologia - En.eswiwiAún no hay calificaciones
- Glomérulo Renal - Wikipedia, La Enciclopedi A LibreDocumento85 páginasGlomérulo Renal - Wikipedia, La Enciclopedi A LibreAngelica SuarezAún no hay calificaciones
- Anatomia Del Sistema CardiacoDocumento8 páginasAnatomia Del Sistema CardiacoLoren RamirezAún no hay calificaciones
- Infeccion Del Tracto UrinarioDocumento35 páginasInfeccion Del Tracto UrinarioRuben Apaza ApazaAún no hay calificaciones
- ValvulopatiasDocumento5 páginasValvulopatiasWuilliam ApaezAún no hay calificaciones
- Trabajo Del Tejido Epitelial....Documento4 páginasTrabajo Del Tejido Epitelial....Maria Castro Cruz0% (1)
- Células EucariótasDocumento37 páginasCélulas EucariótasJoseManuelPrincepAún no hay calificaciones
- MUCOPOLISACARIDOSISDocumento11 páginasMUCOPOLISACARIDOSISTrujillo LievanoAún no hay calificaciones
- Embriologia Sistema Renal Urogenital 14Documento37 páginasEmbriologia Sistema Renal Urogenital 14AnaAguerrevereAún no hay calificaciones
- Pancreas HistologiaDocumento9 páginasPancreas Histologiaandy gomezAún no hay calificaciones
- Mitocondria y VacuolaDocumento22 páginasMitocondria y VacuolaDagoberto Durán100% (1)
- Carcinoma Papilar Del Tiroides, Variante Esclerosante DifusaDocumento23 páginasCarcinoma Papilar Del Tiroides, Variante Esclerosante DifusaAriel Wilson Almendras DazaAún no hay calificaciones
- Genitales ExternosDocumento7 páginasGenitales ExternosLuisCarlosPerezAlvaradoAún no hay calificaciones
- Resumen-Fisiología de La Secreción PancreáticaDocumento3 páginasResumen-Fisiología de La Secreción PancreáticaJose Alfredo HernandezAún no hay calificaciones
- Función y Estructura de La NefronaDocumento9 páginasFunción y Estructura de La NefronaKarina Em El PiAún no hay calificaciones
- La Microcirculación y El Sistema LinfáticoDocumento13 páginasLa Microcirculación y El Sistema LinfáticoHugo Yañez100% (1)
- Bioquimica Metabolismo de La Prostaglandina y ColesterolDocumento9 páginasBioquimica Metabolismo de La Prostaglandina y Colesterolmarce bastidasAún no hay calificaciones
- Protocolo Diagnóstico de TrombofiliaDocumento5 páginasProtocolo Diagnóstico de TrombofiliaMayra Alejandra Prada SerranoAún no hay calificaciones
- Defecto de Tabique Aortico PumonarDocumento12 páginasDefecto de Tabique Aortico PumonarJUAN CESAR RUIZ ESPINOAún no hay calificaciones
- TÉTANOSDocumento99 páginasTÉTANOSJian GalecioAún no hay calificaciones
- Respiracion Celular II PDFDocumento4 páginasRespiracion Celular II PDFvi19per90Aún no hay calificaciones
- Resumen de Patología: TEMA: PULMÓNDocumento42 páginasResumen de Patología: TEMA: PULMÓNCAMILA NICOLE ULLAURI BERREZUETAAún no hay calificaciones
- Leucemia y Cascada de CoagulaciónDocumento23 páginasLeucemia y Cascada de CoagulaciónmispresentacionesAún no hay calificaciones
- Función Renal, Nefrona y Función TubularDocumento32 páginasFunción Renal, Nefrona y Función TubularRafael Vides IrvingAún no hay calificaciones
- Trab Preg 3 Sem BioDocumento13 páginasTrab Preg 3 Sem BioMargaritaty0% (1)
- Membrana Del EritrocitoDocumento33 páginasMembrana Del EritrocitoJulio Rebo100% (1)
- Mapa Conceptual Cirrosis HepaticaDocumento1 páginaMapa Conceptual Cirrosis HepaticaJosé RafaelAún no hay calificaciones
- Introduccion Al MetabolismoDocumento33 páginasIntroduccion Al MetabolismoJosé RafaelAún no hay calificaciones
- Mapa Conceptual Cirrosis HepaticaDocumento1 páginaMapa Conceptual Cirrosis HepaticaJosé RafaelAún no hay calificaciones
- Triptico DrogadiccionDocumento2 páginasTriptico DrogadiccionJosé Rafael100% (3)
- Triptico DrogadiccionDocumento2 páginasTriptico DrogadiccionJosé Rafael100% (3)
- Recomendaciones: A Alimentación SaludableDocumento2 páginasRecomendaciones: A Alimentación SaludableJosé RafaelAún no hay calificaciones
- Triptico ObesidadDocumento3 páginasTriptico ObesidadJosé RafaelAún no hay calificaciones
- HISTERECTOMIADocumento2 páginasHISTERECTOMIAJosé RafaelAún no hay calificaciones
- Lista Cotejo para Ucam 2015Documento8 páginasLista Cotejo para Ucam 2015José RafaelAún no hay calificaciones
- Generalidades de La Cirugía LaparoscópicaDocumento4 páginasGeneralidades de La Cirugía LaparoscópicaJosé RafaelAún no hay calificaciones
- Caracteristicas de La Ciencia en El Siglo XXDocumento7 páginasCaracteristicas de La Ciencia en El Siglo XXJose PrascaAún no hay calificaciones
- GenéticaDocumento19 páginasGenéticaAshley Melody Mauricio tejadaAún no hay calificaciones
- El Texto ArgumentativoDocumento2 páginasEl Texto ArgumentativoELDRICK MENESESAún no hay calificaciones
- BiotoxDocumento4 páginasBiotoxjohn A. yupanqui tomasAún no hay calificaciones
- Resumen Anticoagulantes Banco de SangreDocumento2 páginasResumen Anticoagulantes Banco de SangreKevîn Chámul100% (2)
- Enfermedades en Ojos PDFDocumento9 páginasEnfermedades en Ojos PDFKakatuar KakatuaAún no hay calificaciones
- Apuntes de Sanidad e Higiene PecuariaDocumento5 páginasApuntes de Sanidad e Higiene Pecuariakarina390% (1)
- Tema 1. BACTERIOLOGIADocumento11 páginasTema 1. BACTERIOLOGIASteph VivasAún no hay calificaciones
- Ascaris LumbricoidesDocumento2 páginasAscaris LumbricoidesRoanmy Schol ChalibAún no hay calificaciones
- El Potencial de Sostenibilidad de Los Asentamientos HumanosDocumento199 páginasEl Potencial de Sostenibilidad de Los Asentamientos HumanosMaNuBrenesAún no hay calificaciones
- M14S3AI5Documento5 páginasM14S3AI5manu bb67% (3)
- TFEUDocumento38 páginasTFEUalejandrosantanaAún no hay calificaciones
- 7 GluconeogenesisDocumento24 páginas7 GluconeogenesisFreddyA.SueroAún no hay calificaciones
- Biologia 1Documento41 páginasBiologia 1Viktor McAún no hay calificaciones
- SeminarioDocumento17 páginasSeminarioMarjorie Isabel ApazaAún no hay calificaciones
- Tumor Phyllodes Corea FinalDocumento108 páginasTumor Phyllodes Corea FinalStef Lolez100% (1)
- Konrad LorenzDocumento5 páginasKonrad Lorenzpitigrili100% (1)
- 10 Preguntas Frecuentes Sobre La HomosexualidadDocumento5 páginas10 Preguntas Frecuentes Sobre La HomosexualidadWalter Casco Oliveira SallesAún no hay calificaciones
- Baciloscopia de Esputo y de HansenDocumento32 páginasBaciloscopia de Esputo y de HansenJesica Perez VergaraAún no hay calificaciones
- SEMANA 1 El Origen de La VidaDocumento2 páginasSEMANA 1 El Origen de La VidaEdgar Sánchez CastilloAún no hay calificaciones
- Proyecto Invertebrados (Carta Pokémon + Maqueta + Presentación) 1º Eso Biología y GeologíaDocumento6 páginasProyecto Invertebrados (Carta Pokémon + Maqueta + Presentación) 1º Eso Biología y GeologíaAngel ValparisAún no hay calificaciones
- Guia de Morfologia Celular PDFDocumento6 páginasGuia de Morfologia Celular PDFCopias EthellAún no hay calificaciones
- Poblaciones y ComunidadesDocumento5 páginasPoblaciones y ComunidadesCaroKastenAún no hay calificaciones