También podría gustarte
- UNI2 - Etica Ciencia y Ambiente - RESUMENDocumento7 páginasUNI2 - Etica Ciencia y Ambiente - RESUMENgerardo guzmanAún no hay calificaciones
- Manual para SoñarDocumento8 páginasManual para Soñarchemis4220% (5)
- Respiracion CelularDocumento5 páginasRespiracion CelularSuitsuRohelineAún no hay calificaciones
- Rutas MetabolicasDocumento35 páginasRutas Metabolicassam100% (1)
- GlucólisisDocumento1 páginaGlucólisisDavid100% (1)
- Anhidrasa CarbónicaDocumento19 páginasAnhidrasa CarbónicaAdriana Mendez100% (1)
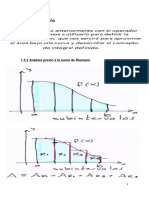
- 1.3calculo IntegralDocumento13 páginas1.3calculo IntegralCarmen Flores0% (1)
- Informe Final Del Proyecto de BETERRAGA - Vasquez Hernandez LeysonDocumento37 páginasInforme Final Del Proyecto de BETERRAGA - Vasquez Hernandez LeysonElianiBazan67% (3)
- BIBLIOGRAFIADocumento4 páginasBIBLIOGRAFIAItalo TrujilloAún no hay calificaciones
- Ciclo KrebsDocumento45 páginasCiclo Krebsandres100% (6)
- GlucolisisDocumento26 páginasGlucolisisAle JoyaAún no hay calificaciones
- Fosforilación OxidativaDocumento8 páginasFosforilación OxidativaLourdes PerdomoAún no hay calificaciones
- El Ciclo de KrebsDocumento16 páginasEl Ciclo de KrebsClemente Peralta Serrano100% (2)
- GlucolisisDocumento20 páginasGlucolisisvaleria de leonAún no hay calificaciones
- GlucogenolisisDocumento15 páginasGlucogenolisisTammy Carrillo Recabarren100% (1)
- Fosforilacion OxidativaDocumento11 páginasFosforilacion OxidativaJose Alberto Ariza PasoAún no hay calificaciones
- Ciclo de KrebsDocumento15 páginasCiclo de KrebsDaniel Saenz DominguezAún no hay calificaciones
- Fosforilacion OxidativaDocumento27 páginasFosforilacion Oxidativajosuekstillo100% (4)
- 6 GluconeogénesisDocumento15 páginas6 Gluconeogénesisdora100% (1)
- Cadena RespiratoriaDocumento2 páginasCadena RespiratoriaJesus RamirezAún no hay calificaciones
- Fosforilacion OxidativaDocumento5 páginasFosforilacion Oxidativaaoihana18100% (1)
- Cadena Respiratoria Exposicion FinalDocumento28 páginasCadena Respiratoria Exposicion Finalesneider alexander perilla vergaraAún no hay calificaciones
- B-Oxidacion y Sintesis de Acidos GrasosDocumento22 páginasB-Oxidacion y Sintesis de Acidos GrasosPatricio Coronado75% (8)
- MicosisDocumento9 páginasMicosisJimy Isaac S-romeroAún no hay calificaciones
- Amortiguadores 2019Documento27 páginasAmortiguadores 2019francisco100% (1)
- Vías de Señalización de ReceptoresDocumento44 páginasVías de Señalización de ReceptoresAngie Alvarez Llanes100% (1)
- GlucolisisDocumento5 páginasGlucolisisDamaris BustamanteAún no hay calificaciones
- Ciclo de KrebsDocumento18 páginasCiclo de Krebscalpaina100% (1)
- Glucogenolisis Glucogénesis Cetogénesis 5Documento45 páginasGlucogenolisis Glucogénesis Cetogénesis 5DianaOrdoñez100% (1)
- METABOLISMO DE LIPÌDOS BioquimicaDocumento33 páginasMETABOLISMO DE LIPÌDOS BioquimicaMartha MoralesAún no hay calificaciones
- Fosforilacion OxidativaDocumento19 páginasFosforilacion OxidativaImanol De la SotaAún no hay calificaciones
- Fosforilación OxidativaDocumento25 páginasFosforilación OxidativaEmilio Jeanpaul Mitacc GutierrezAún no hay calificaciones
- Catabolismo LipidosDocumento34 páginasCatabolismo LipidosJackeline Villanueva SalinasAún no hay calificaciones
- Ciclo de KrebsDocumento33 páginasCiclo de KrebsLN Brenda Janeth Beltrán DíazAún no hay calificaciones
- Sintesis Del ColesterolDocumento2 páginasSintesis Del ColesterolDvid MedranAún no hay calificaciones
- LÍPIDOSDocumento10 páginasLÍPIDOSRafael Angel Huiza Trujillo100% (3)
- CloroplastosDocumento16 páginasCloroplastosclaudioAún no hay calificaciones
- GlucogenolisisDocumento5 páginasGlucogenolisisCristian GonzálezAún no hay calificaciones
- ELECTROLITOSDocumento11 páginasELECTROLITOSMaria Rosario Santolalla Damian100% (2)
- Biosíntesis de Ácidos Grasos y TriglicéridosDocumento46 páginasBiosíntesis de Ácidos Grasos y TriglicéridosXimena Véliz DaudAún no hay calificaciones
- GlucogenolisisDocumento26 páginasGlucogenolisisErnesto LimónAún no hay calificaciones
- ENZIMASDocumento59 páginasENZIMASÉrÌck PÂrédèsAún no hay calificaciones
- GlucólisisDocumento30 páginasGlucólisisJasonRodriguezGuerraAún no hay calificaciones
- GlucogenogenesisDocumento2 páginasGlucogenogenesisGexAún no hay calificaciones
- Rutas MetabolicasDocumento33 páginasRutas MetabolicasJamie Jayjay Martinez100% (2)
- Digestion y Absorcion de ProteinasDocumento3 páginasDigestion y Absorcion de ProteinasYoseimar MarianaAún no hay calificaciones
- Estructura y Función de La HemoglobinaDocumento16 páginasEstructura y Función de La HemoglobinaMiguel Marroquín79% (58)
- Ciclo de KrebsDocumento15 páginasCiclo de KrebsMiguel Angel Rodas Herrera100% (2)
- Vias Metabolicas Trabajo 3.1Documento28 páginasVias Metabolicas Trabajo 3.1Lorgio TR100% (2)
- Ciclo de La Pentosa FosfatoDocumento7 páginasCiclo de La Pentosa Fosfatoeider100% (1)
- Beta OxidacionDocumento20 páginasBeta OxidacionJâzmïn ZårcöAún no hay calificaciones
- Metabolismo y Excrecion de Los FarmacosDocumento27 páginasMetabolismo y Excrecion de Los FarmacosMaria Ines DemeraAún no hay calificaciones
- Ciclo de Krebs InformeDocumento29 páginasCiclo de Krebs InformeyhuverAún no hay calificaciones
- Beta-Oxidacion de Ácidos GrasosDocumento12 páginasBeta-Oxidacion de Ácidos GrasosalezAún no hay calificaciones
- Enfermedad de FarberDocumento5 páginasEnfermedad de FarberDiego CadenaAún no hay calificaciones
- Ensayo LipidosDocumento6 páginasEnsayo Lipidosjhon mueses100% (1)
- CarbohidratosDocumento28 páginasCarbohidratosAnonimo22100% (2)
- GlucolisisDocumento18 páginasGlucolisisManu Camacho100% (2)
- Resumen de Glucólisis y GluconeogénesisDocumento2 páginasResumen de Glucólisis y GluconeogénesisEduardo P. Fernández100% (1)
- Ciclo de KrebsDocumento14 páginasCiclo de KrebsAlexis FonsecaAún no hay calificaciones
- CarbohidratosDocumento8 páginasCarbohidratosAlain MuñozAún no hay calificaciones
- Metabolismo de LípidosDocumento33 páginasMetabolismo de LípidosDavid Alejandro Urbina Meneses100% (1)
- Fosforilacion Oxidativa Teoria QuimiosmoticaDocumento18 páginasFosforilacion Oxidativa Teoria QuimiosmoticaJose Arceo CendejasAún no hay calificaciones
- Ciclo de Krebs MonografiaDocumento23 páginasCiclo de Krebs MonografiaMerelyn Del PiiLar RaMiirez RoMan100% (1)
- Indicadores de Descomposicion Microbiana en Cereales ExpoDocumento14 páginasIndicadores de Descomposicion Microbiana en Cereales ExpoCarmen FloresAún no hay calificaciones
- HaccpDocumento4 páginasHaccpCarmen FloresAún no hay calificaciones
- Determinacion de Humedad en Una Muestra de HarinaDocumento2 páginasDeterminacion de Humedad en Una Muestra de HarinaCarmen FloresAún no hay calificaciones
- Practica PomadaDocumento2 páginasPractica PomadaCarmen FloresAún no hay calificaciones
- Bioconservación IntroduccionDocumento4 páginasBioconservación IntroduccionCarmen FloresAún no hay calificaciones
- Apuntes de Ecuaciones DiferencialesDocumento93 páginasApuntes de Ecuaciones DiferencialesCarmen FloresAún no hay calificaciones
- Requisitos para El Etiquetado de CosméticosDocumento10 páginasRequisitos para El Etiquetado de CosméticosCarmen FloresAún no hay calificaciones
- Microbiología Concepto e HistoriaDocumento23 páginasMicrobiología Concepto e HistoriaKatherine Torres100% (1)
- Estimacion de Riesgos Del Centro Poblado Ollantaytambo PDFDocumento54 páginasEstimacion de Riesgos Del Centro Poblado Ollantaytambo PDFJhoanita AlessaAún no hay calificaciones
- SEP 2 3 Aplicaciones Del MAS MI1909182223Documento2 páginasSEP 2 3 Aplicaciones Del MAS MI1909182223J Melgar MezaAún no hay calificaciones
- CriminalisticaDocumento2 páginasCriminalisticamanuel panchoAún no hay calificaciones
- El Seminario Técnica de Trabajo Académico e InvestigativoDocumento7 páginasEl Seminario Técnica de Trabajo Académico e InvestigativoNicolas Araya CastroAún no hay calificaciones
- Informe Quimica Ambiental Practica N°1, 2, y 3Documento39 páginasInforme Quimica Ambiental Practica N°1, 2, y 3FabianCastilloRodriguez0% (1)
- Plan Anual 2011 Inicial 5 AñosDocumento28 páginasPlan Anual 2011 Inicial 5 Añoschapo222Aún no hay calificaciones
- Ingenieria de YacimientoDocumento63 páginasIngenieria de YacimientoJose Miguel Zambrano FloresAún no hay calificaciones
- 4.2. La Relación Del Empirismo y El Racionalismo Con La Revolución CientíficaDocumento2 páginas4.2. La Relación Del Empirismo y El Racionalismo Con La Revolución CientíficaFernanda Millan Miguez100% (1)
- Parámetros de Crecimiento para MicroorganismosDocumento40 páginasParámetros de Crecimiento para MicroorganismosEli Lozano100% (1)
- Biología TisularDocumento4 páginasBiología TisularGerardo G OsorioAún no hay calificaciones
- Lista Roja Anfibios Del EcuadorDocumento70 páginasLista Roja Anfibios Del EcuadorRobertinoAún no hay calificaciones
- Henry MurrayDocumento7 páginasHenry MurrayRosely de JesúsAún no hay calificaciones
- Practica Domiciliaria - Mecánica de SuelosDocumento2 páginasPractica Domiciliaria - Mecánica de SuelosIngrid Vargas BriceñoAún no hay calificaciones
- Protein AsDocumento16 páginasProtein AsFernando Lopez SanchezAún no hay calificaciones
- Parcial de ErgonomiaDocumento7 páginasParcial de ErgonomiaGisseth Suaza ChaAún no hay calificaciones
- Historia de La PsicologíaDocumento148 páginasHistoria de La PsicologíaBelen Perezlindo100% (1)
- Ensayo Fisica Pelicula Leyes TermodinamicaDocumento3 páginasEnsayo Fisica Pelicula Leyes TermodinamicaGabriel Gonzalez SoleraAún no hay calificaciones
- Royas en ColombiaDocumento20 páginasRoyas en ColombiaJuan David LozanoAún no hay calificaciones
- Mosterin Jesus - Naturaleza Vida Y Cultura PDFDocumento162 páginasMosterin Jesus - Naturaleza Vida Y Cultura PDFcastillaj99100% (9)
- Transporte de Moleculas de Elevada Masa MolecularDocumento1 páginaTransporte de Moleculas de Elevada Masa MolecularMirella CordovaAún no hay calificaciones
- Dicertacion EstructuDocumento12 páginasDicertacion EstructuSergio AlejandroAún no hay calificaciones
- Casas de CampoDocumento6 páginasCasas de CampoLaLho OosuNaAún no hay calificaciones
- CLEGIODocumento9 páginasCLEGIOElioned JR CamaChAún no hay calificaciones
- Geografia Castlla y León para Niños PDFDocumento96 páginasGeografia Castlla y León para Niños PDFÁngel María LaraAún no hay calificaciones
- Los Continentes. Ficha 1. Europa 36. Ficha 2. África 37. Ficha 3. Asia 38. Ficha 4. América 39. Ficha 5. Oceanía 40. Ficha 6 PDFDocumento8 páginasLos Continentes. Ficha 1. Europa 36. Ficha 2. África 37. Ficha 3. Asia 38. Ficha 4. América 39. Ficha 5. Oceanía 40. Ficha 6 PDFcapelloAún no hay calificaciones