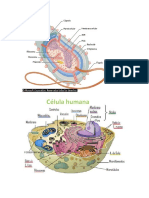
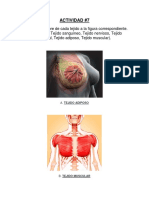
También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Discucion de Bioquimica Bloque III Grupo 1Documento24 páginasDiscucion de Bioquimica Bloque III Grupo 1Monica Amaya75% (4)
- BIOLOGIADocumento11 páginasBIOLOGIAPAULINA SOLIS CORONELAún no hay calificaciones
- Biologia ApuntesDocumento5 páginasBiologia ApuntesAriana RodríguezAún no hay calificaciones
- POLISACARIDODocumento6 páginasPOLISACARIDOGino RodriguezAún no hay calificaciones
- UntitledDocumento36 páginasUntitledJhordy valleAún no hay calificaciones
- Membrana proteínasDocumento3 páginasMembrana proteínasABIGAIL ESPINOZAAún no hay calificaciones
- Membrana CelularDocumento21 páginasMembrana CelularSoledadAún no hay calificaciones
- Investigación PDFDocumento8 páginasInvestigación PDFBautista Reyes Adela RianelAún no hay calificaciones
- La Membrana CelularDocumento46 páginasLa Membrana CelularRandal CGAún no hay calificaciones
- Clase N°10 - Membrana PlasmaticaDocumento20 páginasClase N°10 - Membrana PlasmaticaSergio Esteban GuzmanAún no hay calificaciones
- Membranas Biológicas-Lo EsencialDocumento21 páginasMembranas Biológicas-Lo EsencialElian PerezAún no hay calificaciones
- Copia de 7. LÍPIDOS - EdumedDocumento17 páginasCopia de 7. LÍPIDOS - EdumedCamz NagoAún no hay calificaciones
- Proteínas de La MembranaDocumento8 páginasProteínas de La MembranaMireya Mandujano BendezuAún no hay calificaciones
- Las Membranas de La Célula Ejercen Diversas ActividadesDocumento13 páginasLas Membranas de La Célula Ejercen Diversas ActividadesKauely SouzaAún no hay calificaciones
- Monosacaridos FuncionesDocumento9 páginasMonosacaridos FuncionesISIDRO JONATHAN HERNANDEZ CARMONAAún no hay calificaciones
- Unidad IV. MembranaDocumento57 páginasUnidad IV. Membranamariangeles292003Aún no hay calificaciones
- Seminarios BiologiaDocumento110 páginasSeminarios Biologiapacaesmeralda5Aún no hay calificaciones
- Apuntes Clases Ene Jun 2019Documento121 páginasApuntes Clases Ene Jun 2019Kimberly MartinezAún no hay calificaciones
- Membrana PlasmaticaDocumento14 páginasMembrana PlasmaticaJosefina Cortes100% (2)
- Marco Teorico - CuestionarioDocumento13 páginasMarco Teorico - CuestionarioStephany Mariela Espinoza SachaAún no hay calificaciones
- Taller de MembranasDocumento6 páginasTaller de MembranasMariana Varela AlzateAún no hay calificaciones
- S02C03 - Membrana (I)Documento34 páginasS02C03 - Membrana (I)Alejandro B. SilvaAún no hay calificaciones
- Resumen Biologia Dos DosDocumento17 páginasResumen Biologia Dos DosAilén RestanoAún no hay calificaciones
- La Menbrana Plasmatica-2Documento6 páginasLa Menbrana Plasmatica-2Miau nyanAún no hay calificaciones
- Membrana celular: barrera selectivaDocumento8 páginasMembrana celular: barrera selectivapaudroguettAún no hay calificaciones
- La Celula PDFDocumento49 páginasLa Celula PDFRose NoelAún no hay calificaciones
- Estructura y componentes de la membrana plasmáticaDocumento10 páginasEstructura y componentes de la membrana plasmáticaBryan GomezAún no hay calificaciones
- T ApuntesFC AD2023Documento141 páginasT ApuntesFC AD2023Alondra SerranoAún no hay calificaciones
- Membrana CelularDocumento4 páginasMembrana CelularLuisa Carolina Silva de TuaAún no hay calificaciones
- Membrana plasmática: componentes y funcionesDocumento37 páginasMembrana plasmática: componentes y funcionesAndrés E. Espinoza NolascoAún no hay calificaciones
- Membrana CitoplasmaticaDocumento47 páginasMembrana CitoplasmaticaFernando MartinezAún no hay calificaciones
- Historia de La Teoría de La Membrana Celular 2019Documento33 páginasHistoria de La Teoría de La Membrana Celular 2019JosephDaAún no hay calificaciones
- PROTEÍNASDocumento5 páginasPROTEÍNASFiorelaToAlAún no hay calificaciones
- Sesion 4 PDFDocumento6 páginasSesion 4 PDFAndrés A. Rocha UsugaAún no hay calificaciones
- Capitulo 4 BiocelDocumento12 páginasCapitulo 4 BiocelRENATO ARTURO BAILEY CANTUAún no hay calificaciones
- Membrana Plasmatica 2020-I Mra - S6Documento77 páginasMembrana Plasmatica 2020-I Mra - S6Dino P MoralesAún no hay calificaciones
- Clase No. 06 Solutos Orgánicos e InorganicosDocumento24 páginasClase No. 06 Solutos Orgánicos e InorganicosCamila Ñiquen ParejaAún no hay calificaciones
- 3 Membrana PlasmaticaDocumento30 páginas3 Membrana PlasmaticaSebastian EsnarriagaAún no hay calificaciones
- Proteínas Parte 1 PDFDocumento6 páginasProteínas Parte 1 PDFMichelle GrandeAún no hay calificaciones
- Rol de Las Membranas Biológicas Importancia de Los Receptores y de Las Señales Intracelulares para El Crecimiento y Proliferación CelularDocumento10 páginasRol de Las Membranas Biológicas Importancia de Los Receptores y de Las Señales Intracelulares para El Crecimiento y Proliferación CelularFernan JosimarAún no hay calificaciones
- El OvuloDocumento8 páginasEl OvuloMeimis TorregrozaAún no hay calificaciones
- Membrana CelularDocumento5 páginasMembrana CelularS1A-01-ALVAREZ ZAPATA ARIANA VALENTINAAún no hay calificaciones
- 6 - BiomembranasDocumento17 páginas6 - BiomembranasFlor BovettiAún no hay calificaciones
- Membrana Celular ProyectoDocumento17 páginasMembrana Celular ProyectoGabriel Enrique Giler ReyesAún no hay calificaciones
- Cuestionarios BiofisicaDocumento12 páginasCuestionarios BiofisicaWarismill Peralta AntiguaAún no hay calificaciones
- Membrana celular: estructura y funcionesDocumento37 páginasMembrana celular: estructura y funcionesIdelberto BenitezAún no hay calificaciones
- La Célula PDFDocumento26 páginasLa Célula PDFRuth Alejandra López MaradiagaAún no hay calificaciones
- CelulaDocumento34 páginasCelulaJuan Eduardo Gutierrez TerronesAún no hay calificaciones
- Membrana Celular 3Documento10 páginasMembrana Celular 3JENNIFER ALEXANDRA ARIZA SANDOVALAún no hay calificaciones
- Células y estructura de la membrana plasmáticaDocumento4 páginasCélulas y estructura de la membrana plasmáticaBrenda DominguezAún no hay calificaciones
- Capítulo IV - Adhesión Celular, Uniones Celular y Matriz ExtracelularDocumento12 páginasCapítulo IV - Adhesión Celular, Uniones Celular y Matriz ExtracelularFederico Rivadeneira CornejoAún no hay calificaciones
- Copia de Hidratos, Lipidos y Proteina ResumenDocumento7 páginasCopia de Hidratos, Lipidos y Proteina ResumenLourdes GomezAún no hay calificaciones
- 2 - La Celula y Sus FuncionesDocumento38 páginas2 - La Celula y Sus FuncionesEdi MedAún no hay calificaciones
- Proteinas WienerDocumento40 páginasProteinas WienerRonald franklin balbuenaAún no hay calificaciones
- Estructura de La MembranaDocumento6 páginasEstructura de La MembranaFernando Castro SilvaAún no hay calificaciones
- Informe Re GolgiDocumento7 páginasInforme Re GolgienriqueAún no hay calificaciones
- Las ProteinasDocumento14 páginasLas Proteinasanabely chumacero lizana50% (2)
- Proteinas Integrales de La MembranaDocumento6 páginasProteinas Integrales de La MembranaEduardo Palacios0% (2)
- Resumen Libro Paniagua (Cap 2 Tema 1 y 2)Documento11 páginasResumen Libro Paniagua (Cap 2 Tema 1 y 2)José David Concepción GutiérrezAún no hay calificaciones
- Actividad 4Documento4 páginasActividad 4Sonia MontealegreAún no hay calificaciones
- Transporte celular: fagocitosis, pinocitosis, difusiónDocumento5 páginasTransporte celular: fagocitosis, pinocitosis, difusiónSonia MontealegreAún no hay calificaciones
- Actividad 3Documento18 páginasActividad 3Sonia MontealegreAún no hay calificaciones
- Mapa ConceptualDocumento2 páginasMapa ConceptualSonia MontealegreAún no hay calificaciones
- Actividad 2Documento8 páginasActividad 2Sonia MontealegreAún no hay calificaciones
- La Nutiricion Como Funcion PDFDocumento12 páginasLa Nutiricion Como Funcion PDFSonia MontealegreAún no hay calificaciones
- Actividad 8Documento5 páginasActividad 8Sonia MontealegreAún no hay calificaciones
- Actividad 7Documento3 páginasActividad 7Sonia MontealegreAún no hay calificaciones
- Actividades PreviasDocumento11 páginasActividades PreviasSonia MontealegreAún no hay calificaciones
- SARLAFT-NormasDocumento2 páginasSARLAFT-NormasSonia MontealegreAún no hay calificaciones
- Taller Construyendo Mi EmpresaDocumento5 páginasTaller Construyendo Mi EmpresaSonia MontealegreAún no hay calificaciones
- Taller Construyendo Mi EmpresaDocumento5 páginasTaller Construyendo Mi EmpresaSonia MontealegreAún no hay calificaciones
- Erin BrockovichDocumento5 páginasErin BrockovichBenjamin Alejandro Arcaya NuñezAún no hay calificaciones
- Alteraciones de La Pigmentacion Dermatologia EsteticaDocumento31 páginasAlteraciones de La Pigmentacion Dermatologia EsteticaAmerico I. Ramos MartinezAún no hay calificaciones
- Trastorno Obsesivo-CompulsivoDocumento5 páginasTrastorno Obsesivo-CompulsivoKaren Catari100% (1)
- 5 Oraciones Con Palabras AgudasDocumento5 páginas5 Oraciones Con Palabras AgudasJunior C. Hurtado80% (15)
- Fua Cod 001 - Control CredDocumento18 páginasFua Cod 001 - Control CredUNIDAD DE SEGUROS DIRESA PASCOAún no hay calificaciones
- ChopoDocumento2 páginasChopoAlexia tabarezAún no hay calificaciones
- Examen de Aritmetica-Cuarto Año SecDocumento2 páginasExamen de Aritmetica-Cuarto Año SecJose Paiva MozoAún no hay calificaciones
- Algebraactividadsemana 4Documento9 páginasAlgebraactividadsemana 4Alfredo CoronadoAún no hay calificaciones
- Informe Visita Tecnica Hidroelectrica ManduriacuDocumento9 páginasInforme Visita Tecnica Hidroelectrica ManduriacuKamik LópezAún no hay calificaciones
- Reacciones químicas masa mol leyes ponderalesDocumento6 páginasReacciones químicas masa mol leyes ponderalesOscar RiosAún no hay calificaciones
- Perfil Logistico de China 4Documento13 páginasPerfil Logistico de China 4Julian SilvaAún no hay calificaciones
- ADMINISTRACIÓNDocumento8 páginasADMINISTRACIÓNEli AleAún no hay calificaciones
- AyudaDocumento13 páginasAyudaJhon CondeAún no hay calificaciones
- Guía 1 - Reflexión Individual de La Práctica DocenteDocumento4 páginasGuía 1 - Reflexión Individual de La Práctica DocenteJuan DiegoAún no hay calificaciones
- 1° Cuadernillo VacacionalDocumento18 páginas1° Cuadernillo VacacionalLissette Garcia100% (1)
- Proyecto Flora y FaunaDocumento3 páginasProyecto Flora y FaunaSoledad GaleanoAún no hay calificaciones
- Vender Es Mucho Más. Por Cosimo Chiesa de NegriDocumento15 páginasVender Es Mucho Más. Por Cosimo Chiesa de NegriEdinsonAún no hay calificaciones
- 071-Iroso BaraDocumento8 páginas071-Iroso BaraNiwo PeñalozaAún no hay calificaciones
- Manejo de AlmacenDocumento4 páginasManejo de Almacenurayoy15Aún no hay calificaciones
- Comprobaciones Del Pilar C11Documento54 páginasComprobaciones Del Pilar C11Jose OsorioAún no hay calificaciones
- Costos IIDocumento23 páginasCostos IIJorge ParedesAún no hay calificaciones
- VIH e Implantes DentalesDocumento11 páginasVIH e Implantes DentalesJazmín TortellaAún no hay calificaciones
- Carburantes (1 2)Documento19 páginasCarburantes (1 2)Anai Daniela Gutierrez RiveraAún no hay calificaciones
- Guion de Simulacion de Tecnica AidaDocumento2 páginasGuion de Simulacion de Tecnica Aidalesugamer06Aún no hay calificaciones
- Introducción A La ProgramaciónDocumento6 páginasIntroducción A La ProgramaciónKevin Alvarez Velasquez0% (1)
- Manual Sequax Completo v1.20Documento20 páginasManual Sequax Completo v1.20Enzo UrichAún no hay calificaciones
- 551398-Quintero-GuarínML-y-551385-Sánchez PeñaL-TdGDocumento49 páginas551398-Quintero-GuarínML-y-551385-Sánchez PeñaL-TdGEiner Felipe Mosquera RobledoAún no hay calificaciones
- SecuenciaciónDocumento2 páginasSecuenciaciónleerparasaber100% (1)
- PEUV670707 UV3 FA00000 GHJKLÑDocumento1 páginaPEUV670707 UV3 FA00000 GHJKLÑIvan Emigdio Gonzalez SalazarAún no hay calificaciones