También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Fisiología - Fisiología de PecesDocumento5 páginasFisiología - Fisiología de PecesÑe KsjshdAún no hay calificaciones
- 2.1 Clases de HomeroDocumento34 páginas2.1 Clases de HomeroAndres Ignacio Aguilar AndradeAún no hay calificaciones
- Rta - Celular AdaptativaDocumento5 páginasRta - Celular AdaptativaLucio WekslerAún no hay calificaciones
- Guía Segundo Parcial InmunologíaDocumento7 páginasGuía Segundo Parcial Inmunologíaanaca59505Aún no hay calificaciones
- TCR 1Documento14 páginasTCR 1Kevin alberto Pérez de la cruzAún no hay calificaciones
- Balsas LipidicasDocumento5 páginasBalsas LipidicasJuan Jesus Barrios CapuchinoAún no hay calificaciones
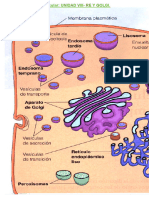
- Distribución y transporte de proteínas en RE, Golgi y lisosomasDocumento5 páginasDistribución y transporte de proteínas en RE, Golgi y lisosomasSara NogueiraAún no hay calificaciones
- Interacciones de las moléculas adrenalina, estrona y colesterolDocumento8 páginasInteracciones de las moléculas adrenalina, estrona y colesterolElizabeth HilarioAún no hay calificaciones
- Estructura molecular membranaDocumento4 páginasEstructura molecular membranaBetsaida Cruz LauraAún no hay calificaciones
- SEMDocumento11 páginasSEMjulissagonzalezzuritaAún no hay calificaciones
- INMUNIDAD CELULAR Estudiantes PDFDocumento49 páginasINMUNIDAD CELULAR Estudiantes PDFlordgimli77100% (3)
- Sistemas de SecreciónDocumento9 páginasSistemas de SecreciónLaura SotoAún no hay calificaciones
- BC Tema 5Documento18 páginasBC Tema 5adfafAún no hay calificaciones
- Composición LipídicaDocumento6 páginasComposición LipídicaZined PMAún no hay calificaciones
- Investigaciones InmunologiaDocumento15 páginasInvestigaciones InmunologiaErick Salazar ManzanoAún no hay calificaciones
- Activacion de Los Linfocitos Clase IIDocumento72 páginasActivacion de Los Linfocitos Clase IIOlga Meléndez MayorquinAún no hay calificaciones
- El retículo endoplasmático y el aparato de GolgiDocumento58 páginasEl retículo endoplasmático y el aparato de GolgiAroa SilvaAún no hay calificaciones
- Retículo Endoplásmico Pi MZVDocumento29 páginasRetículo Endoplásmico Pi MZVCool BoyLVAún no hay calificaciones
- Banco de Preguntas de BiologíaDocumento53 páginasBanco de Preguntas de BiologíaCESAR XAVIER MALDONADO SALCEDOAún no hay calificaciones
- Balsas de membrana y sus implicaciones en procesos celularesDocumento2 páginasBalsas de membrana y sus implicaciones en procesos celularesgerardojr02100% (1)
- Trabajo de BiologiaDocumento11 páginasTrabajo de Biologialoaizaj12Aún no hay calificaciones
- Ayudantía N°4 - Viernes 31.03 - Membrana y NúcleoDocumento41 páginasAyudantía N°4 - Viernes 31.03 - Membrana y NúcleohormilopezAún no hay calificaciones
- Tema 6 AcabadoDocumento13 páginasTema 6 AcabadoMagdalena Oñate SerranoAún no hay calificaciones
- Guia de estudio COMPARTIMIENTOS INTRACELULARESDocumento47 páginasGuia de estudio COMPARTIMIENTOS INTRACELULARESpimenteltamara2Aún no hay calificaciones
- Retículo Endoplasmático RugosoDocumento11 páginasRetículo Endoplasmático RugosoCamila Peralta TamayoAún no hay calificaciones
- Trabajo Tania BiologiaDocumento13 páginasTrabajo Tania BiologiaCarlos Sernaque SaavedraAún no hay calificaciones
- Retículo RugosoDocumento6 páginasRetículo RugosoZuriel HernandezAún no hay calificaciones
- Examen Resuelto Biología Selectividad Junio 2022 AndalucíaDocumento8 páginasExamen Resuelto Biología Selectividad Junio 2022 AndalucíaSonia CalatayudAún no hay calificaciones
- Cuestionario LCTDocumento17 páginasCuestionario LCTMichel Gonzalez100% (1)
- Fichas 7Documento7 páginasFichas 7Noelita RiveraAún no hay calificaciones
- Cap 10Documento7 páginasCap 10Natalia Espíndola BalderasAún no hay calificaciones
- RESPUESTA INMUNE CELULAR CompletaDocumento96 páginasRESPUESTA INMUNE CELULAR CompletaJazmin Sanchez SanchezAún no hay calificaciones
- Clase 7 - Ontogenia de Linfocitos TDocumento10 páginasClase 7 - Ontogenia de Linfocitos TLautaro VasqueAún no hay calificaciones
- Articulo de LipidosDocumento7 páginasArticulo de Lipidosanon-422771100% (1)
- sesiones-2-y-3-organelos-celularesDocumento8 páginassesiones-2-y-3-organelos-celularestik tok videosAún no hay calificaciones
- Capítulo 1Documento21 páginasCapítulo 1John DittelAún no hay calificaciones
- Sistema de Endomembranas IDocumento9 páginasSistema de Endomembranas IRocio RollanAún no hay calificaciones
- Maduración y Activación de Linfocitos TDocumento59 páginasMaduración y Activación de Linfocitos TVioleta Tello MedinaAún no hay calificaciones
- Parcial de Biologia Teoria SamirDocumento4 páginasParcial de Biologia Teoria SamirGeovanis GonzálezAún no hay calificaciones
- Ruta EndociticaDocumento11 páginasRuta EndociticaIsi AnimatedAún no hay calificaciones
- Organelos IIDocumento8 páginasOrganelos IIantonia muñozAún no hay calificaciones
- Tráfico VesicularDocumento55 páginasTráfico VesicularAficionadoAún no hay calificaciones
- Tema 5 Endomembranas Acortado 2018-2019Documento46 páginasTema 5 Endomembranas Acortado 2018-2019Daniel Fernandez-ValenciaAún no hay calificaciones
- Sistemas membranales celulares: Retículo endoplasmáticoDocumento9 páginasSistemas membranales celulares: Retículo endoplasmáticoEnrique Ochoa0% (1)
- El retículo endoplásmico: estructura y función en la célulaDocumento12 páginasEl retículo endoplásmico: estructura y función en la célulaBelén FernándezAún no hay calificaciones
- La Membrana CelularDocumento16 páginasLa Membrana Celularcfabrizzio90Aún no hay calificaciones
- Examen FisioDocumento3 páginasExamen FisioOnis Daniel RodríguezAún no hay calificaciones
- Distribución y Transporte de ProteínasDocumento41 páginasDistribución y Transporte de ProteínasJhonny Ponce Agui100% (2)
- Resolucion Guia 4 - Ayudantias DBIO1070Documento11 páginasResolucion Guia 4 - Ayudantias DBIO1070MarianaAún no hay calificaciones
- B 117 Sistema de EndomembranasDocumento103 páginasB 117 Sistema de EndomembranasGabriel SiqueiraAún no hay calificaciones
- 9° Apunte de Cátedra Biologia Humana Retículo Endoplasmático Y Complejo de Golgi Tráfico VesicularDocumento11 páginas9° Apunte de Cátedra Biologia Humana Retículo Endoplasmático Y Complejo de Golgi Tráfico VesiculardanitzaAún no hay calificaciones
- Membrana Celular Transporte (1) NuevoDocumento21 páginasMembrana Celular Transporte (1) NuevoRodrigo DavalosAún no hay calificaciones
- Biología Re y GolgiDocumento6 páginasBiología Re y GolgiNabila InsfranAún no hay calificaciones
- Organelos PDFDocumento72 páginasOrganelos PDFVictor CarpioAún no hay calificaciones
- Re, Golgi, Lisosoma, Peroxisoma, MitocondriaDocumento118 páginasRe, Golgi, Lisosoma, Peroxisoma, MitocondriaJosselin Saavedra LópezAún no hay calificaciones
- Sistema de EndomembranasDocumento27 páginasSistema de EndomembranasSofía TorresAún no hay calificaciones
- Regulación de La Expresión Génica EucariotasDocumento20 páginasRegulación de La Expresión Génica EucariotasGloribel Bolívar100% (1)
- Inmunidad InnataDocumento22 páginasInmunidad InnataIsabellaAún no hay calificaciones
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- Respuesta inmune adaptativa vs innataDocumento3 páginasRespuesta inmune adaptativa vs innataKarla Bastida100% (1)
- Inmunidad 3Documento30 páginasInmunidad 3MaXiiLovPerGueAún no hay calificaciones
- Resumen HematoDocumento7 páginasResumen HematoManuel VelasquezAún no hay calificaciones
- Hematolgia RecuentosDocumento7 páginasHematolgia RecuentosAlex BenavidesAún no hay calificaciones
- HematologíaDocumento25 páginasHematologíaAndrea Carolina Carrasco RodriguezAún no hay calificaciones
- 8º Guía 6 PERIODO 2Documento15 páginas8º Guía 6 PERIODO 2MARIBELAún no hay calificaciones
- Imnunología Clase 4, 5, 6, 7, 8, 9, 10, 11Documento233 páginasImnunología Clase 4, 5, 6, 7, 8, 9, 10, 11Angie GarciaAún no hay calificaciones
- Materia de Clase InmunoDocumento78 páginasMateria de Clase InmunoBannii RodriguezAún no hay calificaciones
- Guia de Trabajo Biologia - Iv°medio - Sist - InmuneDocumento4 páginasGuia de Trabajo Biologia - Iv°medio - Sist - InmuneRoberto Pacheco100% (1)
- Inmunologia Unidad 1 - USMP 2007Documento49 páginasInmunologia Unidad 1 - USMP 2007TRAFA89Aún no hay calificaciones
- Evaluaciones Pag48 51Documento4 páginasEvaluaciones Pag48 51riveritav0% (1)
- Cap #7 Receptores Inmunitarios y Transducción de SeñalesDocumento92 páginasCap #7 Receptores Inmunitarios y Transducción de SeñalesVILLALVAZO SALVATIERRA VANESSA BELENAún no hay calificaciones
- LeucopoyesisDocumento21 páginasLeucopoyesisbetamerakAún no hay calificaciones
- Trabajo de MetodologiaDocumento7 páginasTrabajo de MetodologiaDumar PerezAún no hay calificaciones
- GLOSARIODocumento6 páginasGLOSARIOCristian CabreraAún no hay calificaciones
- AntígenosDocumento29 páginasAntígenosLuis David Bello RodríguezAún no hay calificaciones
- Cuadernillo de Biología 4 AñoDocumento20 páginasCuadernillo de Biología 4 AñoNoelia RiveroAún no hay calificaciones
- Vih SidaDocumento9 páginasVih SidaPamela JiménezAún no hay calificaciones
- Respuesta inmune adquiridaDocumento14 páginasRespuesta inmune adquiridaAlberto MirandaAún no hay calificaciones
- Sistema Linfatico PDFDocumento44 páginasSistema Linfatico PDFFeer YañezAún no hay calificaciones
- Leucemia Linfocitica CronicaDocumento3 páginasLeucemia Linfocitica Cronicakaiva3Aún no hay calificaciones
- Aparatos Genitales Masculino y FemeninoDocumento18 páginasAparatos Genitales Masculino y Femeninobambull RAún no hay calificaciones
- Material Informativo Session 04Documento2 páginasMaterial Informativo Session 04Yordy CruzAún no hay calificaciones
- Facultad medicina sangreDocumento39 páginasFacultad medicina sangreJose Arias AyalaAún no hay calificaciones
- Enfermedades AutoimunesDocumento17 páginasEnfermedades AutoimunesLiz Estrada100% (1)
- Unidad 7 InfeccionesDocumento8 páginasUnidad 7 InfeccionesEmi GurfinkelAún no hay calificaciones
- Fisiopatologia APUNTE FinalDocumento15 páginasFisiopatologia APUNTE FinalJessie MerinoAún no hay calificaciones
- Introducción A TejidosDocumento37 páginasIntroducción A TejidosGemhma CastellanosAún no hay calificaciones