También podría gustarte
- Act.11 ResumenCap46 AbonceKarisa 301Documento8 páginasAct.11 ResumenCap46 AbonceKarisa 301Andrea AbonceAún no hay calificaciones
- Diferenciales. Cada Tipo Es Muy Sensible A Una Clase de Estímulo Sensitivo para El Que Está Diseñado y Es CasiDocumento5 páginasDiferenciales. Cada Tipo Es Muy Sensible A Una Clase de Estímulo Sensitivo para El Que Está Diseñado y Es CasiJhony PradelaAún no hay calificaciones
- Receptores Sensitivos 2Documento12 páginasReceptores Sensitivos 2kamiila TrujilloAún no hay calificaciones
- Capitulo 47 13° EdiciónDocumento15 páginasCapitulo 47 13° EdiciónAlvaro CarrasquillaAún no hay calificaciones
- Tema 3 - Fisiología General de La SensibilidadDocumento10 páginasTema 3 - Fisiología General de La SensibilidadLaura Villalba EsparzaAún no hay calificaciones
- Receptores sensoriales y procesamiento neuronalDocumento8 páginasReceptores sensoriales y procesamiento neuronalKony Ramos RojasAún no hay calificaciones
- La Información Llega Al Sistema Nervioso Suministrada Por Los Receptores Sensitivos Que Detectan Estímulos de Este TipoDocumento5 páginasLa Información Llega Al Sistema Nervioso Suministrada Por Los Receptores Sensitivos Que Detectan Estímulos de Este TipoYisus GX7Aún no hay calificaciones
- Cap 46 Receptores Sensitivos, Circuitos Neuronales para El Procesamiento de La InformaciónDocumento8 páginasCap 46 Receptores Sensitivos, Circuitos Neuronales para El Procesamiento de La Informaciónrodrigo4590Aún no hay calificaciones
- Receptores Sensoriales, Circuitos Neuronales para Procesando Información PDFDocumento81 páginasReceptores Sensoriales, Circuitos Neuronales para Procesando Información PDFItzel TejedaAún no hay calificaciones
- Guía Expocision FisioDocumento8 páginasGuía Expocision FisioLuz Dary Orbes ConstainAún no hay calificaciones
- Cap 47 Ecpetores Sensitivos y Circuitos NeuronalesDocumento9 páginasCap 47 Ecpetores Sensitivos y Circuitos NeuronalesdavidmoralesAún no hay calificaciones
- CAPÍTULO 47 - Fisiología GUYTONDocumento3 páginasCAPÍTULO 47 - Fisiología GUYTONRashel MoscosoAún no hay calificaciones
- Receptores Sensitivos PT1Documento37 páginasReceptores Sensitivos PT1Cuche1992Aún no hay calificaciones
- Clase 1. AlejandroDocumento14 páginasClase 1. AlejandroMariaJoseMejiasAún no hay calificaciones
- Sistema Sensitivo - Cuestionario 2 FisioloDocumento13 páginasSistema Sensitivo - Cuestionario 2 FisioloEduardo Javier Perez RiosAún no hay calificaciones
- Cap 46 Receptores SensitivosDocumento19 páginasCap 46 Receptores SensitivosQuito TomAún no hay calificaciones
- Resumen Guyton 558-570Documento4 páginasResumen Guyton 558-570Dani ArreguinAún no hay calificaciones
- Circuitos NeuronalesDocumento64 páginasCircuitos NeuronalesKatie BrownAún no hay calificaciones
- Los Receptores SensorialesDocumento14 páginasLos Receptores SensorialesmixoneqAún no hay calificaciones
- Conceptos de Sensación SomáticaDocumento6 páginasConceptos de Sensación SomáticanicolehurtadosAún no hay calificaciones
- Modelo de Informe ChelitaDocumento20 páginasModelo de Informe ChelitaSalomón ChávezAún no hay calificaciones
- 15 Clase - 21-06 - Sistema NerviosoDocumento17 páginas15 Clase - 21-06 - Sistema NerviosoDiana LugoAún no hay calificaciones
- Resumen Capitulo 47Documento7 páginasResumen Capitulo 47Litza Naomi SantosAún no hay calificaciones
- Unidad 4. Guía de Potencial SensorialDocumento4 páginasUnidad 4. Guía de Potencial SensorialGleydel MontañoAún no hay calificaciones
- SINAPSISDocumento7 páginasSINAPSISAnabella VargasAún no hay calificaciones
- Resumen Fisiologia Neuro GuytonDocumento14 páginasResumen Fisiologia Neuro GuytonMiguel BadelAún no hay calificaciones
- Tema 3 NeuroDocumento8 páginasTema 3 NeuroAnabella VargasAún no hay calificaciones
- Sistema somatosensorial y receptores sensorialesDocumento8 páginasSistema somatosensorial y receptores sensorialesAndrea Bulnes MatamorosAún no hay calificaciones
- Tema 6 TextoDocumento14 páginasTema 6 Textoluis marti gomezAún no hay calificaciones
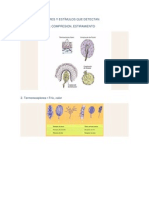
- Tipos de Receptores y Estímulos Que DetectanDocumento10 páginasTipos de Receptores y Estímulos Que DetectanRiichard PiierceAún no hay calificaciones
- Semana 1 Sistema SensorialDocumento12 páginasSemana 1 Sistema SensorialLuis HerediaAún no hay calificaciones
- Receptores SensorialesDocumento6 páginasReceptores SensorialesleonardoAún no hay calificaciones
- Receptores SensorialesDocumento3 páginasReceptores SensorialeslancelotAún no hay calificaciones
- Capítulo 47 Fisiología Guyton y Hall 14 Ed.Documento9 páginasCapítulo 47 Fisiología Guyton y Hall 14 Ed.Manuela MenendezAún no hay calificaciones
- Fisio Sensitivo IIDocumento18 páginasFisio Sensitivo IILeandro Torres CalabiAún no hay calificaciones
- Resumen - Receptores Sensitivos, Circulatorios Neuronales para El Procesamiento de InformacionDocumento11 páginasResumen - Receptores Sensitivos, Circulatorios Neuronales para El Procesamiento de Informacioncarolina caroAún no hay calificaciones
- Relajantes Musculares, E.Documento4 páginasRelajantes Musculares, E.nikholl22Aún no hay calificaciones
- 2.9. RECEPTORES SENSITIVOS, CIRCUITOS NEURONALES PARA EL PROCESAMIENTO Dr. CadimaDocumento26 páginas2.9. RECEPTORES SENSITIVOS, CIRCUITOS NEURONALES PARA EL PROCESAMIENTO Dr. CadimaJair Reynoso Romero100% (1)
- Sistema SomatosensorialDocumento9 páginasSistema Somatosensorialmelody980Aún no hay calificaciones
- Tema 2. Sistema Nervioso SensorialDocumento8 páginasTema 2. Sistema Nervioso SensoriallucisAún no hay calificaciones
- Receptores SensorialesDocumento10 páginasReceptores SensorialesJhuver Alex Campos VasquezAún no hay calificaciones
- Sensibilidad SomaticaDocumento19 páginasSensibilidad Somaticarodrigo ortigaAún no hay calificaciones
- Bases Filosoficas Del Comportamiento Semana 3Documento7 páginasBases Filosoficas Del Comportamiento Semana 3Asumi Panduro SuárezAún no hay calificaciones
- Receptores Sensitivos PDFDocumento56 páginasReceptores Sensitivos PDFPerla LP100% (9)
- Laura FISIO 2 Preguntas OrdenadasDocumento6 páginasLaura FISIO 2 Preguntas OrdenadasPauli GonzalezAún no hay calificaciones
- Receptor EsDocumento11 páginasReceptor EsANDRES SALAZAR NIETOAún no hay calificaciones
- Receptor Sensorial - Wikipedia, La Enciclopedia LibreDocumento27 páginasReceptor Sensorial - Wikipedia, La Enciclopedia LibreEmeron HoriangelizAún no hay calificaciones
- SistemaSomatosensorialDocumento3 páginasSistemaSomatosensorialaleAún no hay calificaciones
- 5-Sistema SomatosensorialDocumento13 páginas5-Sistema SomatosensorialJavier FernandezAún no hay calificaciones
- Cuestionario S.N.C. Emperatriz IngaDocumento9 páginasCuestionario S.N.C. Emperatriz IngaEmperatrizIngaHidalgoAún no hay calificaciones
- PRACTICA NO. 6-Receptores-Sensoriales EQ 5Documento12 páginasPRACTICA NO. 6-Receptores-Sensoriales EQ 5Joguard Lofcraft100% (1)
- Funciones Sensoriales Sistema Somatosensorial PDFDocumento6 páginasFunciones Sensoriales Sistema Somatosensorial PDFOscar CarvajalAún no hay calificaciones
- ReceptoresDocumento3 páginasReceptoresNeida J. Flores R.Aún no hay calificaciones
- Práctica 1 Receptores SensorialesDocumento14 páginasPráctica 1 Receptores SensorialesDaniel James Arriaga80% (5)
- 47 Receptores Sensitivos, Circuitos Neuronales para El Procesamiento de La InformaciónDocumento3 páginas47 Receptores Sensitivos, Circuitos Neuronales para El Procesamiento de La Informaciónileana palaciosAún no hay calificaciones
- Clase #1 - Sistema Nervioso SomaticoDocumento13 páginasClase #1 - Sistema Nervioso SomaticoMariaJoseMejiasAún no hay calificaciones
- Receptores ESTOMADocumento24 páginasReceptores ESTOMANefertiti Akemilo Lola100% (1)
- Fisiología de Los Organos de Los SentidosDocumento16 páginasFisiología de Los Organos de Los Sentidosehtoledoc95% (21)
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- Gen ALKDocumento2 páginasGen ALKGianni AlsinaAún no hay calificaciones
- Formulario de ConsentimientoDocumento1 páginaFormulario de ConsentimientoGianni AlsinaAún no hay calificaciones
- Cap 45 + TresguerresDocumento15 páginasCap 45 + TresguerresGianni AlsinaAún no hay calificaciones
- ReglasMnemotecnicas PDFDocumento72 páginasReglasMnemotecnicas PDFGeorgia Tavara Chorres100% (1)
- Radiologia MusculoesqueleticaDocumento6 páginasRadiologia MusculoesqueleticaGianni AlsinaAún no hay calificaciones
- Bases Epistemologicas PDFDocumento14 páginasBases Epistemologicas PDFpsicotianshiAún no hay calificaciones
- Valvulopatías en geriatríaDocumento5 páginasValvulopatías en geriatríamonica mahechaAún no hay calificaciones
- AdolescenciaDocumento4 páginasAdolescenciaGianni AlsinaAún no hay calificaciones
- Cultura y SociedadDocumento3 páginasCultura y SociedadpaolaAún no hay calificaciones
- Guía Métodos Anticonceptivos y EtsDocumento3 páginasGuía Métodos Anticonceptivos y EtsAiraAún no hay calificaciones
- Autoestima en 10 díasDocumento106 páginasAutoestima en 10 díasJaneth MarquezAún no hay calificaciones
- Ficha Jurisprudencial, Reparación Integral de PerjuiciosDocumento7 páginasFicha Jurisprudencial, Reparación Integral de PerjuiciosanthonygabrielculchacAún no hay calificaciones
- Encuesta 5s - Grupo3Documento3 páginasEncuesta 5s - Grupo3Aileen Paolita Cortez MamaniAún no hay calificaciones
- ReviewDocumento4 páginasReviewKelly MolinaAún no hay calificaciones
- Cocinamos Con María Zannia ThermomixDocumento2 páginasCocinamos Con María Zannia ThermomixCristian BotozisAún no hay calificaciones
- BPM - Laboratorio 9 - MayonesaDocumento15 páginasBPM - Laboratorio 9 - MayonesaCamila CarvalloAún no hay calificaciones
- RSU 3 CASOS CLINICOS GENETICA 1 2023 A LecturaDocumento4 páginasRSU 3 CASOS CLINICOS GENETICA 1 2023 A LecturaJhurney Amilcar TOSCANO CÁCERESAún no hay calificaciones
- Tarea Autonomo BiologiaDocumento13 páginasTarea Autonomo BiologiaMARIA LAPOAún no hay calificaciones
- Autoeficacia - Introducción y MetodologíaDocumento8 páginasAutoeficacia - Introducción y MetodologíaARIANA MARIA OSORIO ALEGRIAAún no hay calificaciones
- Presentacion Precauciones EstándarDocumento34 páginasPresentacion Precauciones EstándarCarolinaAún no hay calificaciones
- ViolenciaDocumento1 páginaViolenciaFernanda Paulina Rodriguez LopezAún no hay calificaciones
- Calculos para El Curso de NutricionDocumento11 páginasCalculos para El Curso de NutricionVictor CastroAún no hay calificaciones
- Check-List-Auditoria AmbientalDocumento3 páginasCheck-List-Auditoria AmbientalWalter Peña89% (9)
- Foucault - El Sentido Histórico de La AlienaciónDocumento15 páginasFoucault - El Sentido Histórico de La AlienaciónAna AlonsoAún no hay calificaciones
- MEDCOMDocumento60 páginasMEDCOMAlejandro MCAún no hay calificaciones
- Crecimiento y Desarrollo Del Sistema EstomatognáticoDocumento20 páginasCrecimiento y Desarrollo Del Sistema EstomatognáticoMarcela Aroca ChandiaAún no hay calificaciones
- Plan de Formacion Talento Humano DAYANAdocxDocumento12 páginasPlan de Formacion Talento Humano DAYANAdocxADRIANA GOMEZ PICOAún no hay calificaciones
- Actividad 13 Importancia de Tener Un PlanDocumento9 páginasActividad 13 Importancia de Tener Un PlanNAYDELI KARIN CARMEN MOREAún no hay calificaciones
- Informe Numerico y Narrativo. BulmaDocumento82 páginasInforme Numerico y Narrativo. BulmaBarron Tenorio VanessaAún no hay calificaciones
- Reaccion de AjusteDocumento4 páginasReaccion de AjusteSederianaAún no hay calificaciones
- Guía de Actividades y Rúbrica de Evaluación - Unidad 2 - Fase 3 - Parámetros Zootécnicos, Instalaciones y Equipos, Nutrición y AlimentaciónDocumento11 páginasGuía de Actividades y Rúbrica de Evaluación - Unidad 2 - Fase 3 - Parámetros Zootécnicos, Instalaciones y Equipos, Nutrición y AlimentaciónYurani RojasAún no hay calificaciones
- Gars 2Documento1 páginaGars 2Aliocha Soraya Ibañez Soto67% (3)
- Guia de Procedimientos de HematologiaDocumento18 páginasGuia de Procedimientos de HematologiaJuan UrbinaAún no hay calificaciones
- Tarea 3 AptitudesDocumento6 páginasTarea 3 AptitudesGerineidy espinal la pazAún no hay calificaciones
- Matriz de Riesgo Publico ExcelDocumento8 páginasMatriz de Riesgo Publico ExcelYurely PinedaAún no hay calificaciones
- Guia Telefonica PUCEDocumento27 páginasGuia Telefonica PUCEVILLARREAL BASKETBALL ACADEMYAún no hay calificaciones
- Plan de Negocios Agencia de Enfermería para Atención DomiciliariaDocumento5 páginasPlan de Negocios Agencia de Enfermería para Atención DomiciliariaRODRIGUEZ PEREZ JOSE EDUARDO0% (1)
- Calcioantagonistas EvipressDocumento3 páginasCalcioantagonistas EvipressGerardo Olivas HernandezAún no hay calificaciones