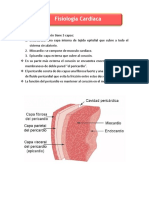
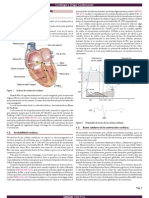
También podría gustarte
- APB01 TareaDocumento5 páginasAPB01 TareaJosimar10 lopez100% (1)
- Biofisica CardíacaDocumento28 páginasBiofisica CardíacaMarcos Días100% (1)
- Fisiologia CardiacaDocumento11 páginasFisiologia CardiacaCarol IsidroAún no hay calificaciones
- Teoria Delm Musculo CardiacoDocumento12 páginasTeoria Delm Musculo CardiacoAlbert KantAún no hay calificaciones
- Electrofisiologia Cardiaca 1Documento9 páginasElectrofisiologia Cardiaca 1Grease Cubas AlzamoraAún no hay calificaciones
- Fisiologia CardiacaDocumento19 páginasFisiologia CardiacaRosa BlancoAún no hay calificaciones
- Clase 3 4-10Documento4 páginasClase 3 4-10Valentina UbillaAún no hay calificaciones
- MIS OBJETIVOS Propiedades Electrofisiológicas Del Miocardio Contráctil y EspecializadoDocumento4 páginasMIS OBJETIVOS Propiedades Electrofisiológicas Del Miocardio Contráctil y EspecializadoPIKAAún no hay calificaciones
- FCV03. Excitación y Conducción CardíacaDocumento37 páginasFCV03. Excitación y Conducción CardíacaSofiaAún no hay calificaciones
- EVA 1 RepasoDocumento3 páginasEVA 1 RepasoCATALINA PIZARRO HERRERAAún no hay calificaciones
- Fisiología Músculo Cardiaco - 3Documento48 páginasFisiología Músculo Cardiaco - 3Daniel Prince TorregrosaAún no hay calificaciones
- Fisiología Del Musculo Cardiaco en Bufo Bufo Como Modelo ExperimentalDocumento6 páginasFisiología Del Musculo Cardiaco en Bufo Bufo Como Modelo ExperimentalDiego Jose Campos LeonAún no hay calificaciones
- ARRITMIASDocumento21 páginasARRITMIASJoselyn RodriguezAún no hay calificaciones
- Electrofisiologia Cardiaca CMDM PDFDocumento10 páginasElectrofisiologia Cardiaca CMDM PDFWendy OrtizAún no hay calificaciones
- Fisiologia Cardiaca GuytonDocumento20 páginasFisiologia Cardiaca GuytonLuis LopezAún no hay calificaciones
- UABP 2. Músculo Cardíaco y Ciclo CardíacoDocumento22 páginasUABP 2. Músculo Cardíaco y Ciclo CardíacoMartín PanicoAún no hay calificaciones
- Revisión de Las Arritmias: Manual MSD Versión para ProfesionalesDocumento11 páginasRevisión de Las Arritmias: Manual MSD Versión para Profesionalesapi-664596295Aún no hay calificaciones
- Fibras CardiacasDocumento5 páginasFibras CardiacasClaudia DiazAún no hay calificaciones
- Musculo Cardiaco Preparatoria EvaDocumento15 páginasMusculo Cardiaco Preparatoria Evanachocoolpro855Aún no hay calificaciones
- Fisiologia CardiacaDocumento28 páginasFisiologia CardiacaDenisse RiosAún no hay calificaciones
- Electrofisiología Cardíaca Material de Exposición Grupo 1Documento13 páginasElectrofisiología Cardíaca Material de Exposición Grupo 1Nicol OrtegaAún no hay calificaciones
- Aparato Circulatorio Fisiología 2023Documento49 páginasAparato Circulatorio Fisiología 2023emmanuelvillaAún no hay calificaciones
- Tema 19 Corazón, FisiologíaDocumento10 páginasTema 19 Corazón, FisiologíaElsa gambraAún no hay calificaciones
- Tema 3.3 FisioDocumento41 páginasTema 3.3 FisioMich VegaAún no hay calificaciones
- Control de La Función CardiovascularDocumento10 páginasControl de La Función CardiovascularMartin Ojeda CarpioAún no hay calificaciones
- Apuntes Del Sistema CardiovascularDocumento23 páginasApuntes Del Sistema CardiovascularAdriana López-ValverdeAún no hay calificaciones
- Cap - 9 (Musculo Cardiaco El Corazon)Documento85 páginasCap - 9 (Musculo Cardiaco El Corazon)JONATHAN ARIEL LUNA JIMENEZAún no hay calificaciones
- Mecanismo Del CorazonDocumento11 páginasMecanismo Del CorazonJc TovarAún no hay calificaciones
- Sistema Cardionector Del CorazónDocumento20 páginasSistema Cardionector Del Corazónalejavanegas011406Aún no hay calificaciones
- Potencial de Membrana y CardiovasculaDocumento7 páginasPotencial de Membrana y CardiovasculaJacki ServianAún no hay calificaciones
- Corazón Musculo Cardiaco ResumenesDocumento16 páginasCorazón Musculo Cardiaco ResumenesLuciana BerriAún no hay calificaciones
- Fisiología CardiacaDocumento12 páginasFisiología CardiacaMiriam Loreley Cahuantzi FloresAún no hay calificaciones
- Desfibrilador CardiacoDocumento46 páginasDesfibrilador CardiacoJesus Resta NavarroAún no hay calificaciones
- Cardiología CTODocumento95 páginasCardiología CTOEfren Penaflores100% (1)
- Clase 4 Fisiologia - Prop Cardiacas y Sist de ConduccionDocumento9 páginasClase 4 Fisiologia - Prop Cardiacas y Sist de ConduccionJaqui De La CruzAún no hay calificaciones
- Arritmias CardiacasDocumento16 páginasArritmias CardiacasJuan Ignacio GuerraAún no hay calificaciones
- Fisiología CardiovascularDocumento19 páginasFisiología CardiovascularGalo CamarenaAún no hay calificaciones
- Sistema CardionectorDocumento10 páginasSistema CardionectorNicolas Bravo JimenezAún no hay calificaciones
- Resumo Fisiologia IIDocumento64 páginasResumo Fisiologia IIRebeca d'El-ReyAún no hay calificaciones
- En El Corazón Las Células Cardiacas Se Dividen en Varios TiposDocumento4 páginasEn El Corazón Las Células Cardiacas Se Dividen en Varios TiposYAIR ESTEBAN ZU�IGA ROSEROAún no hay calificaciones
- Capitulo 9 - Corazon y Contraccion de Musculo CardiacoDocumento4 páginasCapitulo 9 - Corazon y Contraccion de Musculo CardiacoNeilly Valentina Gelvez GarciaAún no hay calificaciones
- Bolillero FINAL - ARTI II CARDIODocumento10 páginasBolillero FINAL - ARTI II CARDIOMedicina UnlamAún no hay calificaciones
- Fisiologia Cardiaca 1Documento93 páginasFisiologia Cardiaca 1soborno5656Aún no hay calificaciones
- Fisiologia Cardiaca GuytonDocumento20 páginasFisiologia Cardiaca Guytonmate la femina100% (1)
- Tipeo Completo Curso ECGDocumento51 páginasTipeo Completo Curso ECGVioleta Canales PueblaAún no hay calificaciones
- Tipeo Semana 1 ECGDocumento10 páginasTipeo Semana 1 ECGVioleta Canales PueblaAún no hay calificaciones
- Preguntero Con Respuestas - CompletoDocumento55 páginasPreguntero Con Respuestas - CompletoFranciscoMolinaAún no hay calificaciones
- Fisiopato ArrtmiasDocumento5 páginasFisiopato ArrtmiasCarla ValenzuelaAún no hay calificaciones
- Introduccion A La Fisiologia CardiacaDocumento16 páginasIntroduccion A La Fisiologia CardiacaMartin PiattiAún no hay calificaciones
- Fármacos Usados en El Tratamiento de Las ArritmiasDocumento13 páginasFármacos Usados en El Tratamiento de Las ArritmiasValentinaAún no hay calificaciones
- Sistema de ConduccionDocumento7 páginasSistema de Conduccionapi-472592451Aún no hay calificaciones
- Sistema CardionectorDocumento36 páginasSistema CardionectorJosé Carlos Pineda100% (8)
- El Corazon Como BombaDocumento9 páginasEl Corazon Como BombaEro Crespo MuñozAún no hay calificaciones
- Fisiologia CardiacaDocumento17 páginasFisiologia CardiacaGenesis PradaAún no hay calificaciones
- Fisiología Cardiovascular Electrocardiografía N°4 PDFDocumento22 páginasFisiología Cardiovascular Electrocardiografía N°4 PDFKevin Argotti CazarAún no hay calificaciones
- Introducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludDe EverandIntroducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludAún no hay calificaciones
- Coagulación Para Todos: Medicina Para TodosDe EverandCoagulación Para Todos: Medicina Para TodosCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Mejorar la Circulación Sanguínea de Forma Natural: Sanar el Sistema Circulatorio y inmunológico para Prevenir Enfermedades, Aumentar la Energía, Aliviar el Estrés y Sentirte Mejor que NuncaDe EverandMejorar la Circulación Sanguínea de Forma Natural: Sanar el Sistema Circulatorio y inmunológico para Prevenir Enfermedades, Aumentar la Energía, Aliviar el Estrés y Sentirte Mejor que NuncaCalificación: 4.5 de 5 estrellas4.5/5 (7)
- Maltrato y Abandono, Relaciones Familiares y Personales Del Adulto MayorDocumento4 páginasMaltrato y Abandono, Relaciones Familiares y Personales Del Adulto MayorMarjorie PérezAún no hay calificaciones
- EnsayoDocumento5 páginasEnsayoMarjorie Pérez75% (4)
- Zoologia ExposicionDocumento3 páginasZoologia ExposicionMarjorie PérezAún no hay calificaciones
- Medula Osea HistologiaDocumento4 páginasMedula Osea HistologiaMarjorie PérezAún no hay calificaciones
- Economia Del EcuadorDocumento5 páginasEconomia Del EcuadorMarjorie PérezAún no hay calificaciones
- Cuadro FungoidesDocumento4 páginasCuadro FungoidesJason HardingAún no hay calificaciones
- Azul BelgaDocumento9 páginasAzul BelgaAlma Maria Mora50% (2)
- Control Calidad de Productos CarnicosDocumento11 páginasControl Calidad de Productos CarnicosDiego Alejandro Rayo VargasAún no hay calificaciones
- 9 Flores para El Alma 2 PDFDocumento7 páginas9 Flores para El Alma 2 PDFJorge TurenneAún no hay calificaciones
- T4 FubicoDocumento9 páginasT4 FubicoBelén Alatriste FortesAún no hay calificaciones
- Sinestesia y PNLDocumento5 páginasSinestesia y PNLbunkeraleAún no hay calificaciones
- Practica 5 AnatomiaDocumento6 páginasPractica 5 AnatomiaYoliii Regidor OlivaAún no hay calificaciones
- Agua y ElectrolitosDocumento32 páginasAgua y ElectrolitosPedro CevallosAún no hay calificaciones
- Clase 7 Sistema NerviosoDocumento35 páginasClase 7 Sistema NerviosoSoleRodriguez100% (1)
- Desnutricion PediatricaDocumento61 páginasDesnutricion PediatricaGerson QuentaAún no hay calificaciones
- Roce de Lineas ArbolesDocumento2 páginasRoce de Lineas ArbolesJuan Andres Espinoza Droguett100% (1)
- Guia 3. Periodo II Sistema Oseo de Los Seres Vivos. Naturales Septimo 2021Documento5 páginasGuia 3. Periodo II Sistema Oseo de Los Seres Vivos. Naturales Septimo 2021rocio menaAún no hay calificaciones
- Radicales Libres, Sistema Del Citocromo P450 y Utilización Del ATPDocumento15 páginasRadicales Libres, Sistema Del Citocromo P450 y Utilización Del ATPKevin Minchola CastañedaAún no hay calificaciones
- Guía - Proceso DigestivoDocumento5 páginasGuía - Proceso DigestivoMargarita QuirozAún no hay calificaciones
- 5 Ejercicios de Gimnasia Cerebral o MentalDocumento6 páginas5 Ejercicios de Gimnasia Cerebral o MentalSALVADORAún no hay calificaciones
- Diapositivas Lesiones Por Armas de FuegoDocumento26 páginasDiapositivas Lesiones Por Armas de FuegoJassonRuizSernaquéAún no hay calificaciones
- Aparato UrinarioDocumento14 páginasAparato UrinarioAlex Murrieta CalixtoAún no hay calificaciones
- OseoDocumento9 páginasOseoAndres JaramilloAún no hay calificaciones
- Preguntas 2° Periodo de Biologà A Grado 6° y 7°.Documento7 páginasPreguntas 2° Periodo de Biologà A Grado 6° y 7°.thalianaAún no hay calificaciones
- Excreción en Los AnimalesDocumento22 páginasExcreción en Los AnimalesEsteban AriasAún no hay calificaciones
- English Phonetics and Phonology PDFDocumento10 páginasEnglish Phonetics and Phonology PDFhuroji17994100% (1)
- Portadas PDFDocumento21 páginasPortadas PDFJoan LopezAún no hay calificaciones
- Bioquimica Metabolica TC LBMDocumento51 páginasBioquimica Metabolica TC LBMzaeta16Aún no hay calificaciones
- Resistencia A La InsulinaDocumento8 páginasResistencia A La InsulinaRicardoAún no hay calificaciones
- TOXICOLOGÍA ResumenDocumento11 páginasTOXICOLOGÍA ResumenJavier RojasAún no hay calificaciones
- Biología CorregidoDocumento11 páginasBiología CorregidoJUAN CARLOS ARRIETA CHAMORROAún no hay calificaciones
- Valoracion de Puerquerio FisiologicoDocumento30 páginasValoracion de Puerquerio FisiologicoAndres Escamilla AlejandroAún no hay calificaciones
- Hoja de Metrado en ExcelDocumento7 páginasHoja de Metrado en ExcelAngello Hernandez PeñaAún no hay calificaciones
- Acerca de Ese Beso de Trish Kerr PDFDocumento71 páginasAcerca de Ese Beso de Trish Kerr PDFMaríaGarcíaAún no hay calificaciones