También podría gustarte
- ANTICOAGULANTESDocumento45 páginasANTICOAGULANTESGloria DariaAún no hay calificaciones
- Empiema PleuralDocumento7 páginasEmpiema PleuralGaleno03Aún no hay calificaciones
- Principales Síndromes Del Sistema UrinarioDocumento9 páginasPrincipales Síndromes Del Sistema UrinarioOscar GutierrezAún no hay calificaciones
- Hematología CompletaDocumento5 páginasHematología Completamayrel21Aún no hay calificaciones
- Qué Es La LeucemiaDocumento10 páginasQué Es La LeucemiaSaul Samame CisnerosAún no hay calificaciones
- Tema 13. Sistema HemolinfaticoDocumento19 páginasTema 13. Sistema HemolinfaticoCarolina Longa100% (1)
- Ciclo de KrebsDocumento33 páginasCiclo de KrebsJosé Álvarez RoblesAún no hay calificaciones
- Acidos - Nucleicos Estructura y FuncionDocumento9 páginasAcidos - Nucleicos Estructura y FuncionArdwin Del PozoAún no hay calificaciones
- Hipertension IntracraneanaDocumento11 páginasHipertension IntracraneanaJavier Colli ReyesAún no hay calificaciones
- Fases de La Coagulacion Sanguinea PDFDocumento2 páginasFases de La Coagulacion Sanguinea PDFRyanAún no hay calificaciones
- La PielDocumento2 páginasLa PielMely FranzottiAún no hay calificaciones
- Enfermedad Hemolítica Del Recién NacidoDocumento66 páginasEnfermedad Hemolítica Del Recién NacidoSilvania Ariza MenaAún no hay calificaciones
- 21 Problema de EnfermeriaDocumento9 páginas21 Problema de EnfermeriaLosReyesDelSaborEjmqAún no hay calificaciones
- Torax Inestable y HemomediastinoDocumento19 páginasTorax Inestable y Hemomediastinobrenda marquezAún no hay calificaciones
- Crisis HipertensivaDocumento64 páginasCrisis HipertensivaEddy' Beltrán AzAún no hay calificaciones
- Tema 8 Virus de La DiarreaDocumento26 páginasTema 8 Virus de La Diarrea'Nenii GarciaAún no hay calificaciones
- Fisiopatología Del ShockDocumento25 páginasFisiopatología Del ShockJuan Carlos Sierra PedrazaAún no hay calificaciones
- Hipertension ArterialDocumento46 páginasHipertension ArterialEdgar Dionicio Quisquinay AlcorAún no hay calificaciones
- LinfomasDocumento30 páginasLinfomasYo MisMo Antonio Gonzalez CabralAún no hay calificaciones
- Fisiologia Renal. Parte IIIDocumento29 páginasFisiologia Renal. Parte IIIAlfonsoPetittoDeGregorioAún no hay calificaciones
- VacutainerDocumento3 páginasVacutainerAhtziri MartinezAún no hay calificaciones
- Regulación de La Presión Arterial Tema 5Documento9 páginasRegulación de La Presión Arterial Tema 5Richard BuendiaAún no hay calificaciones
- Taller DesnutricionDocumento8 páginasTaller DesnutricionTatiana GaleanoAún no hay calificaciones
- Leucemias Agudas y CrónicasDocumento6 páginasLeucemias Agudas y CrónicasEduardo Sánchez de la CruzAún no hay calificaciones
- Cap31 Varices Esofago-GastricasDocumento10 páginasCap31 Varices Esofago-GastricasRobert SuizAún no hay calificaciones
- Seminario - EXAMNES COMPLEMENTARIOS Aparato RespiratorioDocumento63 páginasSeminario - EXAMNES COMPLEMENTARIOS Aparato RespiratorioDaniel BracamonteAún no hay calificaciones
- Leucemia Linfatica Cronica. TricoleucemiaDocumento20 páginasLeucemia Linfatica Cronica. TricoleucemiaAdela Marìa P LAún no hay calificaciones
- Leucemia Linfatica AgudaDocumento10 páginasLeucemia Linfatica AgudaAnonymous GNiOkl3nslAún no hay calificaciones
- Cardiomiopatia Is.Documento7 páginasCardiomiopatia Is.Aleli CarranzaAún no hay calificaciones
- Práctica Número 18. Farmacología Renal. Luna López Luis Miguel.Documento40 páginasPráctica Número 18. Farmacología Renal. Luna López Luis Miguel.L Miguel LunaAún no hay calificaciones
- Medicamentos Más Usados en La Uci CoronariaDocumento44 páginasMedicamentos Más Usados en La Uci CoronariaLiam CárdenasAún no hay calificaciones
- COVID 19 y CorazónDocumento52 páginasCOVID 19 y CorazónSMIBA MedicinaAún no hay calificaciones
- Distribucion de Liquidos CorporalesDocumento2 páginasDistribucion de Liquidos CorporalesCindy MartínezAún no hay calificaciones
- Casos Clínicos de Excitabilidad CelularDocumento16 páginasCasos Clínicos de Excitabilidad CelularDamaris Castillo DuarteAún no hay calificaciones
- Monografias de Enfermedad CerebrovascularDocumento22 páginasMonografias de Enfermedad Cerebrovascularcalber18Aún no hay calificaciones
- Medicamentos CardiovascularesDocumento47 páginasMedicamentos CardiovascularesSandy Burgos CabanillasAún no hay calificaciones
- Radiología de Tórax - J Vargas RomeroDocumento27 páginasRadiología de Tórax - J Vargas RomeroFelipe ThomasAún no hay calificaciones
- Guía de La Practica Clínica Cardiopatía HipertensivaDocumento9 páginasGuía de La Practica Clínica Cardiopatía HipertensivaARIAJAYAún no hay calificaciones
- ExanguinotransfusiónDocumento2 páginasExanguinotransfusiónElena RiveroAún no hay calificaciones
- Pruebas de La Función RenalDocumento29 páginasPruebas de La Función RenalYaritza PinargtAún no hay calificaciones
- Anhidrasa Carbonica Casi CasiDocumento27 páginasAnhidrasa Carbonica Casi CasiRodrigo GomezAún no hay calificaciones
- Enfermedades de Los Leucitos, Ganglios, EtcDocumento80 páginasEnfermedades de Los Leucitos, Ganglios, EtcCarlos LunaAún no hay calificaciones
- Practica .LipoproteinasDocumento10 páginasPractica .LipoproteinasKarla Liszet Gonzalez HernandezAún no hay calificaciones
- Insuficiencia CardíacaDocumento3 páginasInsuficiencia CardíacaThalia BenitoAún no hay calificaciones
- Lupus Eritematoso SistemáticoDocumento12 páginasLupus Eritematoso SistemáticoLuisa Fernanda Campos Briceño100% (1)
- Cuadro Hematico CompletoDocumento19 páginasCuadro Hematico CompletoyancyAún no hay calificaciones
- Analisis Clinicos de Liquidos y Fluidos CorporalesDocumento63 páginasAnalisis Clinicos de Liquidos y Fluidos Corporalesdiania17Aún no hay calificaciones
- Microambiente MedularDocumento11 páginasMicroambiente MedularGido Cliner Quispe FernandezAún no hay calificaciones
- Acidosis y Alcalosis MetabólicaDocumento7 páginasAcidosis y Alcalosis MetabólicadomeAún no hay calificaciones
- Fármacos AntiprotozoariosDocumento10 páginasFármacos AntiprotozoariosRicardoAún no hay calificaciones
- Alteraciones Hematológicas Durante El EmbarazoDocumento29 páginasAlteraciones Hematológicas Durante El EmbarazoAna Luz Muñoz CornejoAún no hay calificaciones
- PancitopeniaDocumento4 páginasPancitopeniaBrigida CAAún no hay calificaciones
- Leishmaniasis Tegumentaria AmericanaDocumento26 páginasLeishmaniasis Tegumentaria Americanadaniel c.Aún no hay calificaciones
- ElectrolitosDocumento6 páginasElectrolitosJobita Yaritza López PinedaAún no hay calificaciones
- Células Madre Hematopoyéticas: Provided by Real Academia Nacional de Farmacia: Portal PublicacionesDocumento23 páginasCélulas Madre Hematopoyéticas: Provided by Real Academia Nacional de Farmacia: Portal PublicacionesNicole Betancourt AlvarezAún no hay calificaciones
- Las Células Madre HematopoyéticasDocumento6 páginasLas Células Madre HematopoyéticasReyna_Maridali_9176Aún no hay calificaciones
- Articulo 2 HematologiaDocumento10 páginasArticulo 2 HematologiaRene Gtz SAún no hay calificaciones
- Celulas MadreDocumento9 páginasCelulas Madrearmanditocote94Aún no hay calificaciones
- Hematopoyesis ArticuloDocumento13 páginasHematopoyesis ArticuloMarlene Sinai Melo100% (1)
- Proteina C Reactiva PDFDocumento1 páginaProteina C Reactiva PDFCarlos Canel EgremiAún no hay calificaciones
- Manual de Primeros Auxilios en El Laboratorio (Gutiérrez)Documento19 páginasManual de Primeros Auxilios en El Laboratorio (Gutiérrez)Carlos Canel EgremiAún no hay calificaciones
- Sopa de PanDocumento2 páginasSopa de PanCarlos Canel EgremiAún no hay calificaciones
- Raices Griegas y Latinas PDFDocumento24 páginasRaices Griegas y Latinas PDFCarlos Canel Egremi67% (3)
- Reacciones FebrilesDocumento3 páginasReacciones FebrilesQUIMICO CLINICO WILLIANS SANCHEZ89% (18)
- PRACTIKADocumento4 páginasPRACTIKACarlos Canel EgremiAún no hay calificaciones
- Tinciones PresentacionDocumento18 páginasTinciones PresentacionCarlos Canel EgremiAún no hay calificaciones
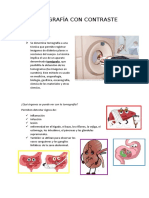
- Tomografía Con ContrasteDocumento7 páginasTomografía Con ContrasteJaneth Rivera100% (5)
- Medicina NuclearDocumento44 páginasMedicina NuclearMauricio Alvarez SalazarAún no hay calificaciones
- Guia de Kit Nutricional Omnilife 2021Documento20 páginasGuia de Kit Nutricional Omnilife 2021CHRISTIAN JAVIER CAGUA VASQUEZAún no hay calificaciones
- Hemorragias de La SegundaDocumento21 páginasHemorragias de La SegundaKarenAún no hay calificaciones
- SEMANA 7 PRACTICA Sedación Consciente TURNO C G2 UNMSMDocumento23 páginasSEMANA 7 PRACTICA Sedación Consciente TURNO C G2 UNMSMJose Ojeda RojasAún no hay calificaciones
- Masaje Con Tratamientos NaturalesDocumento16 páginasMasaje Con Tratamientos NaturalesMiguel RamirezAún no hay calificaciones
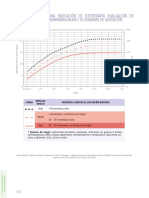
- Curvas para Indicación de FototerapiaDocumento3 páginasCurvas para Indicación de Fototerapiamaria jose NorambuenaAún no hay calificaciones
- Granulopoyesis o GranulocitosDocumento5 páginasGranulopoyesis o GranulocitosAnonymous UnXpJgDi100% (3)
- Estructura de TCC para Insomnio Estudio de CasoDocumento35 páginasEstructura de TCC para Insomnio Estudio de CasoLeonor MoralesAún no hay calificaciones
- Especial FertilidadDocumento8 páginasEspecial FertilidadCecika DanceAún no hay calificaciones
- TESIS COVID 19 Loay NNN (1) ... NMMDocumento43 páginasTESIS COVID 19 Loay NNN (1) ... NMMJorge Joel Salazar MirandaAún no hay calificaciones
- Pruebas Con El DiapazonDocumento2 páginasPruebas Con El Diapazonvianca choquevillcaAún no hay calificaciones
- Ejercicio y Bienestar PsicológicoDocumento21 páginasEjercicio y Bienestar PsicológicoRicardo GFAún no hay calificaciones
- EDITABLE Consentimiento Informado Del Paciente PEDocumento6 páginasEDITABLE Consentimiento Informado Del Paciente PElaboratorio BellidoAún no hay calificaciones
- Hemostasia y TrombosisDocumento28 páginasHemostasia y TrombosisMaría Fernanda Salazar LópezAún no hay calificaciones
- Cuestionario GlomerulopatiasDocumento13 páginasCuestionario GlomerulopatiasNicolle SosaAún no hay calificaciones
- Manual de Primeros Auxilios CUERPO de BOMBEROSDocumento16 páginasManual de Primeros Auxilios CUERPO de BOMBEROSYensy Jose Salazar Serra100% (1)
- Loratadina (Medicamento)Documento8 páginasLoratadina (Medicamento)MARIA FERNANDA OSORIO BOTEROAún no hay calificaciones
- Estudio de Casos Clínicos - NEURO INFANTILDocumento5 páginasEstudio de Casos Clínicos - NEURO INFANTILLoreto Andrea IbarAún no hay calificaciones
- Depresion y AnsiedadDocumento16 páginasDepresion y AnsiedadKattya Idiarte100% (1)
- 3329 pdf-1106915199Documento2 páginas3329 pdf-1106915199Eddy SantallaAún no hay calificaciones
- Tapp y TepDocumento4 páginasTapp y TepkateAún no hay calificaciones
- Derecho de PeticionDocumento5 páginasDerecho de PeticionDaniel PinedaAún no hay calificaciones
- Induccion y ConduccionDocumento45 páginasInduccion y ConduccionElizabeth PinelAún no hay calificaciones
- Epilepsia en OdontologiaDocumento15 páginasEpilepsia en OdontologiaDoris Gómez RubioAún no hay calificaciones
- Lectura Critica 1Documento4 páginasLectura Critica 1Giancarlo Pintado MedinaAún no hay calificaciones
- Caso Clínico S2 PARTODocumento19 páginasCaso Clínico S2 PARTOMaraSifuentesAún no hay calificaciones
- Contusion Pulmonar.2Documento17 páginasContusion Pulmonar.2ed uardoAún no hay calificaciones
- Reh - G - 34 Guia Alteraciones de La ManoDocumento10 páginasReh - G - 34 Guia Alteraciones de La ManoHeimer ErazoAún no hay calificaciones
- Caso Clínico 2 Bolivia (1) ResueltoDocumento3 páginasCaso Clínico 2 Bolivia (1) ResueltoMoises Guarachi SirpaAún no hay calificaciones