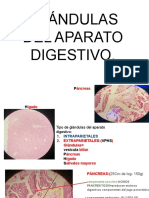
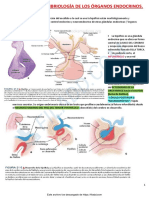
También podría gustarte
- Glándula Tiroides, Paratiroides, Suprarrenales y Páncreas EndócrinoDocumento15 páginasGlándula Tiroides, Paratiroides, Suprarrenales y Páncreas EndócrinoErika LoyarteAún no hay calificaciones
- Consolidacion DefDocumento28 páginasConsolidacion DefLuis Carlos HernandezAún no hay calificaciones
- COAGULACION Y HEMOSTASIA - AsdDocumento10 páginasCOAGULACION Y HEMOSTASIA - AsdBethania Davila100% (1)
- EVC Isquémico FisiopatologiaDocumento58 páginasEVC Isquémico FisiopatologiaSalvador FigueroaAún no hay calificaciones
- Sindrome de Condensación PulmonarDocumento6 páginasSindrome de Condensación PulmonarJuliannys BriceñoAún no hay calificaciones
- Artrosis Artritis ColumnaDocumento64 páginasArtrosis Artritis ColumnaGastónMauricioAún no hay calificaciones
- Fractura de JonesDocumento4 páginasFractura de JonesDanielAún no hay calificaciones
- Histologia Hígado y PáncreasDocumento8 páginasHistologia Hígado y PáncreasJhon VargasAún no hay calificaciones
- Insuficiencia VascularDocumento3 páginasInsuficiencia VascularJose Rafael UmbriaAún no hay calificaciones
- Hepatitis Por MedicamentosDocumento32 páginasHepatitis Por Medicamentos1424robpaz100% (1)
- Sist. Digestivo IiDocumento68 páginasSist. Digestivo IiSabina MoyaAún no hay calificaciones
- Anatomía Del Aparato Genital Femenino - Fernando Vázquez RiveraDocumento59 páginasAnatomía Del Aparato Genital Femenino - Fernando Vázquez RiveraErika Guerrero100% (1)
- Artritis SépticaDocumento17 páginasArtritis SépticaJazmin Brito CastilloAún no hay calificaciones
- CodoDocumento11 páginasCodoMelito AucapiñaAún no hay calificaciones
- Síndromes RestrictivosDocumento32 páginasSíndromes RestrictivosFernando Lopez MartinezAún no hay calificaciones
- Cintura EscapularDocumento8 páginasCintura EscapularDuzzan Patricio Dunn Torres100% (2)
- Hemostasia SecundariaDocumento11 páginasHemostasia SecundariaMartina CortesAún no hay calificaciones
- SRAADocumento5 páginasSRAALuis Chicmana100% (2)
- Presentacion AbdomenDocumento52 páginasPresentacion Abdomenapi-362469425Aún no hay calificaciones
- Esplenopatias QuirurgicasDocumento27 páginasEsplenopatias Quirurgicasapi-26188232100% (2)
- Drenaje PosturalDocumento11 páginasDrenaje PosturalAS ASAún no hay calificaciones
- Distrofia Muscular de Emery - DreifussDocumento14 páginasDistrofia Muscular de Emery - DreifussgeorgeAún no hay calificaciones
- SD Guillian Barre KinesiologiaDocumento34 páginasSD Guillian Barre KinesiologiaGonzalo Andrés Meza CruzAún no hay calificaciones
- EspondiloartropatíasDocumento27 páginasEspondiloartropatíasEliseo ZepedaAún no hay calificaciones
- ARTROPATIASDocumento97 páginasARTROPATIAScared99Aún no hay calificaciones
- Fracturas Del PieDocumento29 páginasFracturas Del Pieyakodude100% (3)
- Protocolo para Dolor Abdominal Agudo ActualDocumento13 páginasProtocolo para Dolor Abdominal Agudo ActualDavid Parco HerreraAún no hay calificaciones
- Incontinencia UrinariaDocumento22 páginasIncontinencia UrinariaWilmer MolinaAún no hay calificaciones
- Anatomía de Cadera (YO)Documento19 páginasAnatomía de Cadera (YO)Gonzalo Martinez SolerAún no hay calificaciones
- Fracturas de La Extremidad InferiorDocumento80 páginasFracturas de La Extremidad InferiorAlfredo PerezAún no hay calificaciones
- Higiene PosturalDocumento2 páginasHigiene PosturalCitas HDMCRAún no hay calificaciones
- Fisiología ColumnaDocumento7 páginasFisiología ColumnaJoselin BastidasAún no hay calificaciones
- Generalidades y Bases de OncologíaDocumento5 páginasGeneralidades y Bases de Oncologíaisidora canalesAún no hay calificaciones
- Práctica 2Documento8 páginasPráctica 2Letty MendezAún no hay calificaciones
- Sindrome ExtrapiramidalDocumento46 páginasSindrome ExtrapiramidalVictor Hernandez100% (1)
- Exploración Física Del AbdomenDocumento54 páginasExploración Física Del AbdomencazaresAún no hay calificaciones
- Deslizameinto EpifisiarioDocumento11 páginasDeslizameinto EpifisiarioMARIANA GOMEZ CASTELLANOSAún no hay calificaciones
- Insuficiencia Vascular PerifericaDocumento62 páginasInsuficiencia Vascular PerifericaMel Palacios RAún no hay calificaciones
- Pancreatitis NecrotizanteDocumento6 páginasPancreatitis NecrotizanteJovanni Alexandre Macias MatusAún no hay calificaciones
- Complicaciones en El EmbarazoDocumento35 páginasComplicaciones en El EmbarazoAndy BalboaAún no hay calificaciones
- Semiología Del Aparato Locomotor FinalDocumento62 páginasSemiología Del Aparato Locomotor FinalJose Bernedo CornejoAún no hay calificaciones
- Benefisios de La Actividad FisicaDocumento10 páginasBenefisios de La Actividad FisicaJonnathan Andy AgamaAún no hay calificaciones
- Estenosis Aortica Severa CriteriosDocumento6 páginasEstenosis Aortica Severa CriteriosSol LakosAún no hay calificaciones
- Síndromes HemorrágicosDocumento57 páginasSíndromes HemorrágicosRoberto ArellanoAún no hay calificaciones
- Sarcoma SinovialDocumento10 páginasSarcoma SinovialEdwin GioAún no hay calificaciones
- Potasio Hipo Hiperkalemia 111123180338 Phpapp01Documento21 páginasPotasio Hipo Hiperkalemia 111123180338 Phpapp01Tania GuisselleAún no hay calificaciones
- Tratamiento Fisioterapéutico para El Paciente DiabéticoDocumento15 páginasTratamiento Fisioterapéutico para El Paciente DiabéticoEdgar Alejandro Salinas EliasAún no hay calificaciones
- Valvulopatia Mitral - Estenosis e InsuficienciaDocumento15 páginasValvulopatia Mitral - Estenosis e InsuficienciaAndrés DuqueAún no hay calificaciones
- Aparato Reproductor Femenino 8Documento25 páginasAparato Reproductor Femenino 8MARIA CELESTE COLQUE ARCE100% (1)
- Síndromes PleuropulmonaresDocumento16 páginasSíndromes PleuropulmonarestsukikagesamaAún no hay calificaciones
- Embriologia e Histologia de EndocrinoDocumento11 páginasEmbriologia e Histologia de EndocrinoJessica R.L 18Aún no hay calificaciones
- Farmacología de Anestésicos LocalesDocumento58 páginasFarmacología de Anestésicos LocalesMartin SernaAún no hay calificaciones
- Masas Abdominales. DaddyDocumento104 páginasMasas Abdominales. DaddySakura la bonita100% (1)
- TM4 TraumatismoRaquimedularDocumento31 páginasTM4 TraumatismoRaquimedularIgnacio Rodriguez HeviaAún no hay calificaciones
- Sistema Venoso SuperficialDocumento6 páginasSistema Venoso SuperficialAlejandro RomeroAún no hay calificaciones
- Enfermedad Inflamatoria Intestinal. PresentaciónDocumento33 páginasEnfermedad Inflamatoria Intestinal. PresentaciónAnand Prakash KaurAún no hay calificaciones
- TromboflebitisDocumento2 páginasTromboflebitisCamilo CortesAún no hay calificaciones
- Sistema Renina AngiotensinaDocumento8 páginasSistema Renina AngiotensinaIgor TorrezAún no hay calificaciones
- Procesos Administrativos SaludDocumento84 páginasProcesos Administrativos SaludAleliz ZCAún no hay calificaciones
- FX RotulaDocumento20 páginasFX RotulaAleliz ZC100% (2)
- Plateado de VidrioDocumento3 páginasPlateado de VidrioAleliz ZCAún no hay calificaciones
- 7 Cadena EpidemiológicaDocumento25 páginas7 Cadena EpidemiológicaAleliz ZCAún no hay calificaciones
- Plateado de VidrioDocumento3 páginasPlateado de VidrioAleliz ZCAún no hay calificaciones
- Como Utilizar Las VelasDocumento14 páginasComo Utilizar Las VelasSonia Montiel100% (1)
- Hoja de TiempoDocumento31 páginasHoja de TiempoEudo Gonzalez0% (1)
- Tres Leyes de NewtonDocumento2 páginasTres Leyes de NewtonemanuelAún no hay calificaciones
- Alisado BrasilenoDocumento3 páginasAlisado BrasilenoLOLI1250% (2)
- Fichas Técnicas de Lubricantes y Bombas de Gasolina de Ciertos AceitesDocumento9 páginasFichas Técnicas de Lubricantes y Bombas de Gasolina de Ciertos AceitesAlex VidalAún no hay calificaciones
- DaniloDocumento36 páginasDaniloLuis Gavino AnlasAún no hay calificaciones
- La Ciencia Moderna Conceptos y CaracterísticasDocumento1 páginaLa Ciencia Moderna Conceptos y CaracterísticasThania ValeraAún no hay calificaciones
- Fase 4 - Reconociendo Los Enfoques PsicológicosDocumento8 páginasFase 4 - Reconociendo Los Enfoques PsicológicosLauraAún no hay calificaciones
- Informe 2Documento3 páginasInforme 2Sara JimenexAún no hay calificaciones
- Escritos Literarios-Arthur SchopenhauerDocumento108 páginasEscritos Literarios-Arthur SchopenhauerSebastián D'UrsiAún no hay calificaciones
- El BuqueDocumento9 páginasEl BuqueJulio JoseAún no hay calificaciones
- Factores de Los Riesgos - Zenobio, KenjoDocumento5 páginasFactores de Los Riesgos - Zenobio, KenjoKENJO FABRICIO ZENOBIO CASTILLOAún no hay calificaciones
- Grado 3°, P II - Indagadores CIENCIASDocumento3 páginasGrado 3°, P II - Indagadores CIENCIASSandra CalderonAún no hay calificaciones
- FuerzaDocumento138 páginasFuerzaPaul Y RubyAún no hay calificaciones
- Tecnicas Especiales HistoquimicaDocumento41 páginasTecnicas Especiales HistoquimicaAzís Humberto Gastelum ZatarainAún no hay calificaciones
- Ecología Alimentaria de Ucides Occidentalis Del Sector Occidental de La Bahía JiquiliscoDocumento13 páginasEcología Alimentaria de Ucides Occidentalis Del Sector Occidental de La Bahía JiquiliscoCarlos RiveraAún no hay calificaciones
- Charles Leiter - Bautismo o CristoDocumento12 páginasCharles Leiter - Bautismo o CristoBenjamin A. FigueroaAún no hay calificaciones
- Caso Pizza Inc Grupo 2Documento10 páginasCaso Pizza Inc Grupo 2julio correaAún no hay calificaciones
- Maranon eDocumento295 páginasMaranon eLeslie CastilloAún no hay calificaciones
- La Tradición Culinaria China Goza de Popularidad MundialDocumento5 páginasLa Tradición Culinaria China Goza de Popularidad MundialRita RolónAún no hay calificaciones
- Formulario - Ecuaciones Diferenciales PDFDocumento3 páginasFormulario - Ecuaciones Diferenciales PDFJose RamirezAún no hay calificaciones
- Catálogo Volta Motors Perú 2022v 05.08.22Documento22 páginasCatálogo Volta Motors Perú 2022v 05.08.22Gamers ProAún no hay calificaciones
- Etica para La VidaDocumento9 páginasEtica para La VidaEdgar CamargosAún no hay calificaciones
- QuimmicaDocumento26 páginasQuimmicaAdrian C LermoAún no hay calificaciones
- San Basilio Magno-Atento A Ti MismoDocumento11 páginasSan Basilio Magno-Atento A Ti MismocarobmAún no hay calificaciones
- Corolario, Lemas, Ley, Teorema, Postulado, DoctrinaDocumento2 páginasCorolario, Lemas, Ley, Teorema, Postulado, DoctrinaCintia Rialtamirano0% (1)
- AROMATERAPIADocumento2 páginasAROMATERAPIAHeyra SalinasAún no hay calificaciones
- Proyecto Comida Shuar - 122138Documento15 páginasProyecto Comida Shuar - 122138Paul Quintanilla0% (1)
- Scrubber: Manual de InstruccionesDocumento52 páginasScrubber: Manual de InstruccionesHenry FonsecaAún no hay calificaciones
- Analisis de Estructuras Semestre I - 2010 Certamen 3 PROBLEMA 1 (50 Puntos)Documento8 páginasAnalisis de Estructuras Semestre I - 2010 Certamen 3 PROBLEMA 1 (50 Puntos)Jake SHAún no hay calificaciones