También podría gustarte
- Cascada EnzimaticaDocumento5 páginasCascada EnzimaticaDaniela VargasAún no hay calificaciones
- Factores de CoagulaciónDocumento7 páginasFactores de CoagulaciónErnesto DavidAún no hay calificaciones
- Cascada de La CoagulaciónDocumento11 páginasCascada de La Coagulaciónfernando olmos paredesAún no hay calificaciones
- Factores de CoagulaciónDocumento16 páginasFactores de CoagulaciónAndrés ContrerasAún no hay calificaciones
- Vias de CoagulacionDocumento3 páginasVias de CoagulacionAlvaro MejiaAún no hay calificaciones
- Monografia Cascada de Coagulacion PresentacionDocumento20 páginasMonografia Cascada de Coagulacion PresentacionJohnxito AjotaAún no hay calificaciones
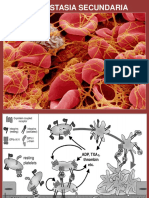
- Hemostasia Secundaria - Pruebas de La Hemostasia 28834 Andressa Dornelas Da Silva SantanaDocumento12 páginasHemostasia Secundaria - Pruebas de La Hemostasia 28834 Andressa Dornelas Da Silva SantanaAcassio VianaAún no hay calificaciones
- Cascada de Coagulación. BORRADORDocumento3 páginasCascada de Coagulación. BORRADORKarla Fernanda Ruiz BáezAún no hay calificaciones
- Fisiologia de La Hemostasia y Coagulacion SanguineaDocumento6 páginasFisiologia de La Hemostasia y Coagulacion SanguineaYong MondragonAún no hay calificaciones
- Coagulación - Wikipedia, La Enciclopedia LibreDocumento101 páginasCoagulación - Wikipedia, La Enciclopedia LibreLuisa LópezAún no hay calificaciones
- Fisiología de La CoagulacionDocumento7 páginasFisiología de La CoagulacionGaby GallegosAún no hay calificaciones
- Cascada de CoagulacionDocumento9 páginasCascada de Coagulacionsacg2435Aún no hay calificaciones
- Seminario. HemostasiaDocumento10 páginasSeminario. Hemostasiaarios_409594100% (1)
- Objetivos 9 y 10 Discu FisioDocumento4 páginasObjetivos 9 y 10 Discu FisioEric CalderónAún no hay calificaciones
- Cascada de CoagulaciónDocumento18 páginasCascada de CoagulaciónFernando MedinaAún no hay calificaciones
- Coagulacion Sanguinea-Grupo Sanguineos 2Documento75 páginasCoagulacion Sanguinea-Grupo Sanguineos 2heyitsadry2112Aún no hay calificaciones
- P1C5 Clase BIIDocumento50 páginasP1C5 Clase BIIÁngel Alejandro Galdàmez GarciaAún no hay calificaciones
- CoagulaciónDocumento18 páginasCoagulaciónAnildibethAún no hay calificaciones
- CoagulaciónDocumento19 páginasCoagulaciónRazo MateoAún no hay calificaciones
- Hemostasia y CoagulaciónDocumento29 páginasHemostasia y CoagulaciónMendoza Rodríguez Danna MichelleAún no hay calificaciones
- Fase PlasmáticaDocumento5 páginasFase PlasmáticaIgnacio Zahid Osuna SosaAún no hay calificaciones
- Hemostasia, Hemorragia Quirúrgica y TransfusiónDocumento12 páginasHemostasia, Hemorragia Quirúrgica y TransfusiónMario SalazarAún no hay calificaciones
- La Cascada de CoagulaciónDocumento10 páginasLa Cascada de CoagulaciónCinthia Chalaco100% (1)
- Investigación CoagulaciónDocumento2 páginasInvestigación Coagulaciónalejandra ZepedaAún no hay calificaciones
- Cascada de Coagulación - Luis Fernando Guzmán GuzmánDocumento7 páginasCascada de Coagulación - Luis Fernando Guzmán GuzmánLuis Fernando Guzman GuzmanAún no hay calificaciones
- HEMOSTASIA PRIMARIA Secundaria, FibrinolisisDocumento3 páginasHEMOSTASIA PRIMARIA Secundaria, Fibrinolisisalexandra montalvo0% (1)
- Factor de CoagulaciónDocumento19 páginasFactor de CoagulaciónMario OliveraAún no hay calificaciones
- HemostasiaDocumento2 páginasHemostasiaKarina Laura MastroleoAún no hay calificaciones
- Presentacion. COAGULACION.2Documento20 páginasPresentacion. COAGULACION.2Janacua Figueroa DianaAún no hay calificaciones
- Informe HEMOSTASIA Alan GutierrezDocumento9 páginasInforme HEMOSTASIA Alan Gutierrezeduardoivanhurtadochavez20Aún no hay calificaciones
- Cascada de La Coagulacion y Pruebas TP y TTPDocumento9 páginasCascada de La Coagulacion y Pruebas TP y TTPLiiz VazquezAún no hay calificaciones
- Fisiología TermodinámicaDocumento4 páginasFisiología TermodinámicaSilvanaDiazAún no hay calificaciones
- Clase N°2 Factores, Mecanismos y Vías de La CoagulaciónDocumento11 páginasClase N°2 Factores, Mecanismos y Vías de La CoagulaciónDelia Heather Molina GarridoAún no hay calificaciones
- Sistema de La PlasminaDocumento2 páginasSistema de La PlasminaAlejandro PvAún no hay calificaciones
- Cascada de La CoagulacxiónDocumento5 páginasCascada de La Coagulacxióngabicita2002Aún no hay calificaciones
- CoagulaciónDocumento21 páginasCoagulaciónYuderkis Moreno RamirezAún no hay calificaciones
- T4 - Ciru - Hemostasia y Transfusion SanguineaDocumento14 páginasT4 - Ciru - Hemostasia y Transfusion SanguineaPablo Segales BautistaAún no hay calificaciones
- Hemostasia Secundaria NC Feb 13Documento47 páginasHemostasia Secundaria NC Feb 13JosePallaresGarnica100% (1)
- Hemostasia, Coagulacion, Fibrinolisis y AnticougulantesDocumento15 páginasHemostasia, Coagulacion, Fibrinolisis y AnticougulantesEvelynn GarcíaAún no hay calificaciones
- Monografia FisiologiaDocumento25 páginasMonografia Fisiologiajonathan alexander salinas calderonAún no hay calificaciones
- Cascada CoagulacionDocumento13 páginasCascada CoagulacionDiego PintoAún no hay calificaciones
- Fisiopatología de Los Trastornos de La Coagulación 2007Documento16 páginasFisiopatología de Los Trastornos de La Coagulación 2007api-3742102100% (5)
- Mecanismos de La HemostasiaDocumento18 páginasMecanismos de La HemostasiaAndres PelayoAún no hay calificaciones
- Vía Extrínseca e Intrínseca de La CoagulaciónDocumento3 páginasVía Extrínseca e Intrínseca de La Coagulaciónwilliam ramosAún no hay calificaciones
- Las Cascadas de CoagulaciónDocumento3 páginasLas Cascadas de CoagulaciónAlaska YoungAún no hay calificaciones
- El Sistema de La Coagulación Está Integrado Por Una Serie de Proteínas PlasmáticasDocumento3 páginasEl Sistema de La Coagulación Está Integrado Por Una Serie de Proteínas Plasmáticasuo23220029Aún no hay calificaciones
- Teà Rico Coagulacià NDocumento13 páginasTeà Rico Coagulacià NMartina LOPEZ RIVOIRAAún no hay calificaciones
- Clase Uss Fisiologia de La HemostasiaDocumento88 páginasClase Uss Fisiologia de La HemostasiaBerni BreyerAún no hay calificaciones
- Bloque 5 Parte 1Documento8 páginasBloque 5 Parte 1laura marina melero aanchezAún no hay calificaciones
- Cascada de Coagulación ClásicaDocumento11 páginasCascada de Coagulación ClásicaRobi GeltAún no hay calificaciones
- Hemostasia SecundariaDocumento26 páginasHemostasia SecundariaMonserrat Duarte MunozAún no hay calificaciones
- La Fisiología de La HemostasiaDocumento21 páginasLa Fisiología de La Hemostasiayuli andrea pinto pabaAún no hay calificaciones
- Cascada de CoagulaciónDocumento13 páginasCascada de CoagulaciónDaniela SandovalAún no hay calificaciones
- Cascada de CoagulaciónDocumento3 páginasCascada de CoagulaciónCaper RH50% (2)
- ANTITROMBÓTICOSDocumento33 páginasANTITROMBÓTICOSLore y Juan InglésAún no hay calificaciones
- Fármacos Anticoagulantes, Tombolíticos y AntiplaquetariosDocumento13 páginasFármacos Anticoagulantes, Tombolíticos y AntiplaquetariosJaidaly Andreina Avila MedinaAún no hay calificaciones
- Coagulación Para Todos: Medicina Para TodosDe EverandCoagulación Para Todos: Medicina Para TodosCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- Rejuvenecer Con El Plasma Sanguíneo De Los JóvenesDe EverandRejuvenecer Con El Plasma Sanguíneo De Los JóvenesCalificación: 5 de 5 estrellas5/5 (1)
- pracTICA N9Documento26 páginaspracTICA N9Ernesto GarciaAún no hay calificaciones
- Garcia Vasquez Ernesto - INFORME N°2Documento16 páginasGarcia Vasquez Ernesto - INFORME N°2Ernesto GarciaAún no hay calificaciones
- Farmacologia Irnfome USMPDocumento27 páginasFarmacologia Irnfome USMPErnesto GarciaAún no hay calificaciones
- Fisiología Hepática MARCO TEORICO PARTE 1Documento2 páginasFisiología Hepática MARCO TEORICO PARTE 1Ernesto Garcia100% (1)
- Practica n1Documento18 páginasPractica n1Ernesto GarciaAún no hay calificaciones
- Gluconeogenesis 1Documento34 páginasGluconeogenesis 1Ernesto GarciaAún no hay calificaciones
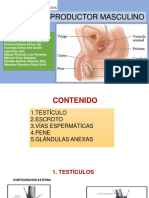
- Aparato Repdroductor MasculinoDocumento40 páginasAparato Repdroductor MasculinoErnesto GarciaAún no hay calificaciones
- Tema 1 - El Origen de La Vida Terrestre ModernaDocumento16 páginasTema 1 - El Origen de La Vida Terrestre ModernaErnesto GarciaAún no hay calificaciones
- Pruebas de CoagulaciónDocumento2 páginasPruebas de CoagulaciónErnesto GarciaAún no hay calificaciones
- Fosas Nasales, Senos Paranasales 2017Documento38 páginasFosas Nasales, Senos Paranasales 2017Ernesto Garcia100% (1)
- Histología Del EstómagoDocumento6 páginasHistología Del EstómagoErnesto GarciaAún no hay calificaciones
- Histologia Del Intestino DelgadoDocumento5 páginasHistologia Del Intestino DelgadoErnesto GarciaAún no hay calificaciones
- Informe, Recuento LeucocitarioDocumento15 páginasInforme, Recuento LeucocitarioStephanieAún no hay calificaciones
- Los Efectos de La Ansiedad en El CerebroDocumento5 páginasLos Efectos de La Ansiedad en El CerebroMax Andre Zapata PazoAún no hay calificaciones
- TurkDocumento1 páginaTurkanon-649429100% (4)
- Modelo 1Documento24 páginasModelo 1Paula LeAún no hay calificaciones
- PolimiositisDocumento7 páginasPolimiositisapi-3698281100% (2)
- Enfermedad Coronaria en Hombres y MujeresDocumento6 páginasEnfermedad Coronaria en Hombres y MujeresDuncan MaxAún no hay calificaciones
- BarotrauDocumento11 páginasBarotrauOlem OsnolaAún no hay calificaciones
- Enfermedad Hemolitica Del Recien NacidoDocumento41 páginasEnfermedad Hemolitica Del Recien NacidoBeto Ramírez100% (2)
- Caracterización de Phlebodium Spp.Documento8 páginasCaracterización de Phlebodium Spp.David GarcíaAún no hay calificaciones
- Maniobra de ValsalvaDocumento6 páginasManiobra de ValsalvajackcisnerosAún no hay calificaciones
- Trabajo Practico Botanica Economica.Documento9 páginasTrabajo Practico Botanica Economica.Andrea Ruiz50% (2)
- Neurologia Pediatrica Clinica PDFDocumento406 páginasNeurologia Pediatrica Clinica PDFAriel Montero50% (2)
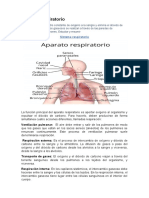
- Sistema RespiratorioDocumento4 páginasSistema RespiratorioCandy De valdezAún no hay calificaciones
- Ic en PediatriaDocumento50 páginasIc en Pediatriacarlos ubillusAún no hay calificaciones
- Palpacion Del SacroDocumento9 páginasPalpacion Del SacroMatices Art PeruAún no hay calificaciones
- 11.-Jeopardy BasesDocumento73 páginas11.-Jeopardy BasesArmando RodríguezAún no hay calificaciones
- Ingreso Eutm 2020Documento3 páginasIngreso Eutm 2020Mauricio VianaAún no hay calificaciones
- Ficha InformativaDocumento7 páginasFicha InformativaCarmen Fajardo ZapataAún no hay calificaciones
- Practica N°2 Llenado de OdontogramaDocumento22 páginasPractica N°2 Llenado de OdontogramaDarwin Reyes ParionaAún no hay calificaciones
- Uworld Notas 2016 PDFDocumento82 páginasUworld Notas 2016 PDFEthar LoveAún no hay calificaciones
- Glosario SSDocumento3 páginasGlosario SSemilioAún no hay calificaciones
- Lipidos 2daDocumento34 páginasLipidos 2daMichelle HuitzAún no hay calificaciones
- Regulación Molecular de La Inducción de La Cresta NeuralDocumento3 páginasRegulación Molecular de La Inducción de La Cresta NeuralNelson SotoAún no hay calificaciones
- Capitulo 5 PDFDocumento41 páginasCapitulo 5 PDFJULIAAún no hay calificaciones
- 10 Tipos de DrogasDocumento4 páginas10 Tipos de DrogasLopez DanyAún no hay calificaciones
- FONÉTICADocumento8 páginasFONÉTICAJesus VargasAún no hay calificaciones
- Instructivo-Para-Registro-De-Productos-Aditivos Agrocalidad PDFDocumento7 páginasInstructivo-Para-Registro-De-Productos-Aditivos Agrocalidad PDFTanabeth del SaltoAún no hay calificaciones
- 4to Informe (Visceras de Pollo)Documento6 páginas4to Informe (Visceras de Pollo)Eduardo Antonio Molinari NovoaAún no hay calificaciones
- HUESOSDocumento8 páginasHUESOSGLADYSAún no hay calificaciones
- Emociones y Sistema Inmune, PPT Autor: Dr. Orestes García FigueiredoDocumento61 páginasEmociones y Sistema Inmune, PPT Autor: Dr. Orestes García FigueiredoNancySáezAún no hay calificaciones