También podría gustarte
- Fichas 3. Sistema Nervioso, Siapsis y Estructura, Dos Fichas de Parafrasis y Una de ResumenDocumento7 páginasFichas 3. Sistema Nervioso, Siapsis y Estructura, Dos Fichas de Parafrasis y Una de ResumenIrving SanchezAún no hay calificaciones
- Artículo Transtorno Obs-CompulsivoDocumento5 páginasArtículo Transtorno Obs-Compulsivodavid19751980Aún no hay calificaciones
- Histologia Sistema Nervioso ContenidoDocumento19 páginasHistologia Sistema Nervioso Contenidovictoria sofiaAún no hay calificaciones
- Unidad IV. Historia de La PsicologiaDocumento10 páginasUnidad IV. Historia de La PsicologiaLuis Enrique Nuñez MinayaAún no hay calificaciones
- Presion ArterialDocumento10 páginasPresion ArterialAnonymous ZIAIJFDfFAún no hay calificaciones
- NeurologíaDocumento120 páginasNeurologíacarlosAún no hay calificaciones
- Regula C I On Autonomic A CardiovascularDocumento36 páginasRegula C I On Autonomic A CardiovascularStephany BruigetAún no hay calificaciones
- Historia de La NeuropsicologíaDocumento6 páginasHistoria de La NeuropsicologíaDaniela AzocarAún no hay calificaciones
- Via AuditivaDocumento11 páginasVia AuditivaPaula O.R100% (1)
- Informe de Exposición PsicofisiologíaDocumento22 páginasInforme de Exposición PsicofisiologíaAndrea MejiaAún no hay calificaciones
- Genética MatematicaDocumento10 páginasGenética MatematicaroyAún no hay calificaciones
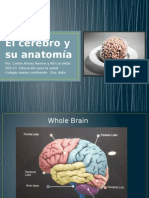
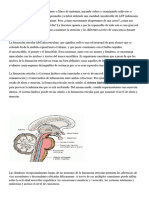
- El Cerebro y Su AnatomíaDocumento20 páginasEl Cerebro y Su Anatomíaimanolordiales50% (2)
- Relatoria Bulbo RaquideoDocumento10 páginasRelatoria Bulbo RaquideoJuanita GarzónAún no hay calificaciones
- Guia 1 GenerneuroanatoDocumento28 páginasGuia 1 GenerneuroanatoBerenice TorresAún no hay calificaciones
- LABORATORIODocumento6 páginasLABORATORIOJohanna OspinaAún no hay calificaciones
- Tálamo InternoDocumento8 páginasTálamo Internogdnm77Aún no hay calificaciones
- Tema 1. Introducción Al Estudio Del Sistema NerviosoDocumento77 páginasTema 1. Introducción Al Estudio Del Sistema NerviosoDarío BejaranoAún no hay calificaciones
- Neuroanatomía-Curso Online PUCDocumento120 páginasNeuroanatomía-Curso Online PUCLauraguilar100% (8)
- Anatomía y Embriología de Órganos y SistemasDocumento167 páginasAnatomía y Embriología de Órganos y SistemasCristóbal Muñoz Pérez100% (2)
- Cerebro PecesDocumento18 páginasCerebro PecesDumar PerezAún no hay calificaciones
- Formacion ReticularDocumento11 páginasFormacion Reticularvargassahori30Aún no hay calificaciones
- PRACTICA No. 1 GENERALIDADES DEL SISTEMA NERVIOSO Dr. Luis Arturo Juárez Islas.Documento8 páginasPRACTICA No. 1 GENERALIDADES DEL SISTEMA NERVIOSO Dr. Luis Arturo Juárez Islas.Julio Perez67% (3)
- Bases Biologicas Del Comportamiento Humano CuestionarioDocumento2 páginasBases Biologicas Del Comportamiento Humano CuestionarioHosswe100% (1)
- Cordon Neural - Fisiología AnimalDocumento13 páginasCordon Neural - Fisiología AnimalSein SaggAún no hay calificaciones
- Argumento Sobre NeurocienciaDocumento8 páginasArgumento Sobre Neurociencianelson anglas castroAún no hay calificaciones
- M1T13 - NeuropscDocumento3 páginasM1T13 - NeuropscThonny EspinosaAún no hay calificaciones
- Orientaciones Metodológicas. Morfo IDocumento11 páginasOrientaciones Metodológicas. Morfo IJavier Diaz CastroAún no hay calificaciones
- Lectura 1Documento5 páginasLectura 1Rocio Caballero CaballeroAún no hay calificaciones
- NEUROFISIOLOGIADocumento175 páginasNEUROFISIOLOGIADavid PaniaguaAún no hay calificaciones
- Neurociencia 4Documento38 páginasNeurociencia 4Luis CahuanaAún no hay calificaciones
- Anato FisioDocumento10 páginasAnato FisioAndrés MardonesAún no hay calificaciones
- NEUROPSICOLOGIA. Caso Practico-Pablo Ayala.Documento27 páginasNEUROPSICOLOGIA. Caso Practico-Pablo Ayala.Psicologia Uac Valdivia UacAún no hay calificaciones
- Sistema Nervioso Periférico. Plexo Braquial y LumbosacroDocumento9 páginasSistema Nervioso Periférico. Plexo Braquial y LumbosacroJulio César Amarilla100% (1)
- Sustancia Reticular - Bases Biol..Documento8 páginasSustancia Reticular - Bases Biol..Vega Bermudez CesarAún no hay calificaciones
- ENSAYODocumento2 páginasENSAYOjulieth100% (1)
- Clase 1 Desarrollo Histórico de La Neurociencia PDFDocumento33 páginasClase 1 Desarrollo Histórico de La Neurociencia PDFjhonAún no hay calificaciones
- Sistema Nervioso EntericoDocumento14 páginasSistema Nervioso EntericoRosyLeyvaAún no hay calificaciones
- Trab cELESTEDocumento16 páginasTrab cELESTEIsabelAún no hay calificaciones
- Astrocitoma de Bajo Grado Visualizado Por Resonancia Magnética Cerebral Con Contraste en Paciente Masculino de 24 AnosDocumento14 páginasAstrocitoma de Bajo Grado Visualizado Por Resonancia Magnética Cerebral Con Contraste en Paciente Masculino de 24 AnosFrancisco ChaconAún no hay calificaciones
- Cap 1 - Elementos Propedéuticos de Neuropsicologia PDFDocumento19 páginasCap 1 - Elementos Propedéuticos de Neuropsicologia PDFKILAMAZUKEAún no hay calificaciones
- Neuropatología TumoralDocumento51 páginasNeuropatología Tumoralapi-3742102100% (5)
- Contenido Científico Neurobiología PDF 2022Documento13 páginasContenido Científico Neurobiología PDF 2022Melany PatriciaAún no hay calificaciones
- Afasias SubcorticalesDocumento12 páginasAfasias SubcorticalesPablo Garza BarreraAún no hay calificaciones
- Presentacion Sistema NerviosoDocumento18 páginasPresentacion Sistema NerviosoAsh bvAún no hay calificaciones
- Capitulo 2 Diojani SepulvedaDocumento7 páginasCapitulo 2 Diojani SepulvedaDiojani Sepulveda Terrero100% (1)
- Diviana Rocca Seccion B Neurofisiologia y Fisiopatologia Unidad 3Documento114 páginasDiviana Rocca Seccion B Neurofisiologia y Fisiopatologia Unidad 3oriana roccaAún no hay calificaciones
- Lenguaje SNCDocumento47 páginasLenguaje SNCmagalyflor100% (1)
- Cap2. Neurociencia y AprendizajeDocumento19 páginasCap2. Neurociencia y AprendizajePaola QuisbertAún no hay calificaciones
- Tarea 4 Historia de La PsicologiaDocumento5 páginasTarea 4 Historia de La PsicologiaignacioAún no hay calificaciones
- Atlas Bases Biológicas Del ComportamientoDocumento41 páginasAtlas Bases Biológicas Del ComportamientoJulio Rodolfo Vasquez Roberts100% (7)
- Resumen Final Neuro FerreDocumento22 páginasResumen Final Neuro FerreMili NunesAún no hay calificaciones
- Unidad 1 La Neurología y Neuropatología Ficha Catedra UnoDocumento15 páginasUnidad 1 La Neurología y Neuropatología Ficha Catedra UnoRocio Perez BonahoraAún no hay calificaciones
- #3 Tallo CerebralDocumento11 páginas#3 Tallo CerebralMonserrat Olivo TalaveraAún no hay calificaciones
- Anatomía Del Sistema Nervioso Central y Periférico Modificadas2Documento63 páginasAnatomía Del Sistema Nervioso Central y Periférico Modificadas2Georgina GalindoAún no hay calificaciones
- Sistema NerviosoDocumento48 páginasSistema NerviosoMiguel Angel Vázquez PérezAún no hay calificaciones
- Tarea 1 AnatomiaDocumento4 páginasTarea 1 Anatomiayamilex DavidAún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- El cannabis en patologías del sistema nervioso centralDe EverandEl cannabis en patologías del sistema nervioso centralAún no hay calificaciones
- Emprendimiento en El Ecuador-Septimo A Grupo 1Documento7 páginasEmprendimiento en El Ecuador-Septimo A Grupo 1Solange RiofrioAún no hay calificaciones
- Plan de Aula # 1 Grado 10 17-03-2020Documento11 páginasPlan de Aula # 1 Grado 10 17-03-2020Diego AleanAún no hay calificaciones
- Tabla de Indicadores CTE 1 Oct-21Documento1 páginaTabla de Indicadores CTE 1 Oct-21Nico RAún no hay calificaciones
- Folleto Actividad 4Documento3 páginasFolleto Actividad 4DENNIS100% (1)
- Pilotaje CS y LyL 2021Documento6 páginasPilotaje CS y LyL 2021Monica CalvaAún no hay calificaciones
- HUAMANI HUAMANI - MergedDocumento8 páginasHUAMANI HUAMANI - MergedShirley GryAún no hay calificaciones
- S03-Dimensión Social La Musica 3 4 5Documento3 páginasS03-Dimensión Social La Musica 3 4 5Joao Aquino100% (1)
- Formacioin Humana I Trabajo FinalDocumento4 páginasFormacioin Humana I Trabajo FinalJosé SamirAún no hay calificaciones
- Problemas de AprendisajeDocumento8 páginasProblemas de AprendisajeAndrea CarolinaAún no hay calificaciones
- CuadroDocumento3 páginasCuadroJAZHIEL CORPUS VALEROAún no hay calificaciones
- Actividades Simulador PDFDocumento1 páginaActividades Simulador PDFCindy GorocicaAún no hay calificaciones
- Decreto 2888 de 2007Documento46 páginasDecreto 2888 de 2007rymendozap100% (1)
- CV RomeliaDocumento2 páginasCV RomeliaEdduMedinaChumaceroAún no hay calificaciones
- Aviso Publicitario Psicopedagogia MiguelDocumento1 páginaAviso Publicitario Psicopedagogia MiguelMIGUEL HERNANDEZAún no hay calificaciones
- Proyecto EducativofedericoDocumento18 páginasProyecto EducativofedericoMarvin AlvaradoAún no hay calificaciones
- Informe EHBADocumento4 páginasInforme EHBADiego PorrasAún no hay calificaciones
- Plan 2001Documento32 páginasPlan 2001غارسيا بيجايسكوسا راكيل اليخاندراAún no hay calificaciones
- Mapa Conceptual - Ciencia, Técnica y TecnologíaDocumento3 páginasMapa Conceptual - Ciencia, Técnica y TecnologíaAnyel door100% (4)
- Actualización Fichas Tecnicas Agosto 2021Documento5 páginasActualización Fichas Tecnicas Agosto 2021Miguel SanabriaAún no hay calificaciones
- Cultura y Contracultura - Diferencias y Similitudes - Cuadro ComparativoDocumento8 páginasCultura y Contracultura - Diferencias y Similitudes - Cuadro ComparativoLuis Felipe Sanchez PeñaAún no hay calificaciones
- Agentes Educativos GRUPO 1Documento6 páginasAgentes Educativos GRUPO 1NILSER JHONY FLORES RODRIGUEZAún no hay calificaciones
- Trastornos de La Conducta AlimentariaDocumento26 páginasTrastornos de La Conducta AlimentariaHerssel Ciro Huamanquispe Tunque100% (1)
- Anexo 7 Semana Del 20 Al 24 de Marzo 2023Documento7 páginasAnexo 7 Semana Del 20 Al 24 de Marzo 2023Johanna NarváezAún no hay calificaciones
- Desarrollo Del Lenguaje ATP M1L3Documento4 páginasDesarrollo Del Lenguaje ATP M1L3Dulce Maria Escobar SeguraAún no hay calificaciones
- Hoja de Trabajo # 1 Actividad # 2 Reglamento Del AprendizDocumento3 páginasHoja de Trabajo # 1 Actividad # 2 Reglamento Del AprendizLEIDY ZAPATAAún no hay calificaciones
- Sesión - Guerra Entre Huascar y AtahualpaDocumento9 páginasSesión - Guerra Entre Huascar y AtahualpaMarcos David Dominguez SampiAún no hay calificaciones
- 019 FormatonGuiande Aprendizaje Medios de Transmision ACT 2Documento12 páginas019 FormatonGuiande Aprendizaje Medios de Transmision ACT 2Diego Ferney Farfán VelásquezAún no hay calificaciones
- Induccion y Capacitacion de PersonalDocumento4 páginasInduccion y Capacitacion de Personalemmy JhoannaAún no hay calificaciones
- Sesión A y C. Dibujo San Jacinto Fe, Costumbres TradicionesDocumento6 páginasSesión A y C. Dibujo San Jacinto Fe, Costumbres TradicionesRogelio Isla RamosAún no hay calificaciones
- Cuadros de Doble EntradaDocumento4 páginasCuadros de Doble EntradaFrida zumiko Vizquerra CañameroAún no hay calificaciones