También podría gustarte
- Absorción con filtro BK como terapia eficaz en el riñón del mieloma múltipleDe EverandAbsorción con filtro BK como terapia eficaz en el riñón del mieloma múltipleAún no hay calificaciones
- Producción y Purificación Parcial de Enzimas Hidrolíticas de Aspergillus Ficuum en Fermentación Sóli PDFDocumento14 páginasProducción y Purificación Parcial de Enzimas Hidrolíticas de Aspergillus Ficuum en Fermentación Sóli PDFriiAún no hay calificaciones
- Radiologia Pediatrica AMPapDocumento107 páginasRadiologia Pediatrica AMPapPaula Milla Torres100% (1)
- Anestesicos GeneralesDocumento7 páginasAnestesicos GeneralesIsaacAún no hay calificaciones
- Farmacologia Basica y Clinica Katzung 14a Edicion OPIOIDESDocumento21 páginasFarmacologia Basica y Clinica Katzung 14a Edicion OPIOIDESAnnery PaulinoAún no hay calificaciones
- Manejo Nutricional Perro PDFDocumento7 páginasManejo Nutricional Perro PDFLuis Carlos VegaAún no hay calificaciones
- 1º Prd. Ap. Ortodoncia y Férulas Oclusales - Curso 2021-2022Documento63 páginas1º Prd. Ap. Ortodoncia y Férulas Oclusales - Curso 2021-2022maríaAún no hay calificaciones
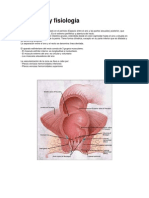
- Anatomía y FisiologíaDocumento12 páginasAnatomía y FisiologíalylipiliAún no hay calificaciones
- Ejercicios de DilucionDocumento6 páginasEjercicios de DilucionKatherine de jesus0% (1)
- RT Costa Rica.Documento8 páginasRT Costa Rica.paoAún no hay calificaciones
- Ix Exploración Del Nervio GolosofaringeoDocumento2 páginasIx Exploración Del Nervio GolosofaringeoMohamed Bilal AbouAún no hay calificaciones
- Estudio de Casos y ControlesDocumento5 páginasEstudio de Casos y ControlesMelanie Fernanda BastidasAún no hay calificaciones
- Metodos de InmunodiagnosticoDocumento5 páginasMetodos de InmunodiagnosticoHarlen Mendoza100% (1)
- Receptores Del EquilibrioDocumento3 páginasReceptores Del EquilibriojimenaGutierrez50% (2)
- Equipos y Accesorios en RadiodiagnósticoDocumento27 páginasEquipos y Accesorios en RadiodiagnósticoMICHEL CAROLINA RODRIGUEZ100% (1)
- Integración MetabólicaDocumento51 páginasIntegración MetabólicaDiego NolascoAún no hay calificaciones
- Al ContrasteDocumento129 páginasAl ContrasteAleXis Neciosup PerezAún no hay calificaciones
- Resumen de Temas UrologiaDocumento134 páginasResumen de Temas UrologiaAlexander CedeñoAún no hay calificaciones
- 2 Farmacologia Basica y Clinica Katzung 14a EdicionDocumento27 páginas2 Farmacologia Basica y Clinica Katzung 14a EdicionMaría Jiménez CaicedoAún no hay calificaciones
- AntimaláricosDocumento21 páginasAntimaláricosJhonatan Efraín López CarbajalAún no hay calificaciones
- Efectos Biologicos de Las RadiacionesDocumento6 páginasEfectos Biologicos de Las Radiacionesannmary2207Aún no hay calificaciones
- Fisiopatologia de La InflamaciónDocumento13 páginasFisiopatologia de La InflamaciónRicardo Ortiz NovilloAún no hay calificaciones
- Factores de Exposición Óptimos de Radiación en Radiología Convencional Y Digital para Obtener Imágenes Diagnosticas de CalidadDocumento4 páginasFactores de Exposición Óptimos de Radiación en Radiología Convencional Y Digital para Obtener Imágenes Diagnosticas de CalidadKaren Franco100% (1)
- Efectos Tardios de La RadiacionDocumento19 páginasEfectos Tardios de La RadiacionRafael Rguez. DíazAún no hay calificaciones
- Normas de Bioseguridad en Medicina NuclearDocumento20 páginasNormas de Bioseguridad en Medicina NuclearMichell CaballeroAún no hay calificaciones
- RibozimasDocumento3 páginasRibozimasSalvador Salcedo67% (3)
- Cuadro AntineoplasicosDocumento3 páginasCuadro AntineoplasicosclaudiogutierrezAún no hay calificaciones
- Imagenologia PedrosaDocumento48 páginasImagenologia PedrosaJ CanAún no hay calificaciones
- Informe SC ParasitosisDocumento28 páginasInforme SC ParasitosisEnder Alvarez Alvarado100% (1)
- ReceptoresDocumento14 páginasReceptoresMauricio Espinoza MaechelAún no hay calificaciones
- 6 Radiologia PDFDocumento15 páginas6 Radiologia PDFWilly Daniel Cipriano CastroAún no hay calificaciones
- Manual de Bioseguridad en Salud Visual y OcularDocumento98 páginasManual de Bioseguridad en Salud Visual y OcularCamila QuevedoAún no hay calificaciones
- Artro MuñecaDocumento9 páginasArtro MuñecaJosé Moya100% (1)
- Rayos X INSTRUCTIVO TRABAJO 4 CATEDRA en Periodo de Gestación.Documento3 páginasRayos X INSTRUCTIVO TRABAJO 4 CATEDRA en Periodo de Gestación.angieAún no hay calificaciones
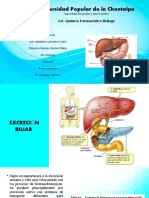
- Excreción BiliarDocumento10 páginasExcreción BiliarFranx SanchezAún no hay calificaciones
- Caso Clinico GiardiaDocumento2 páginasCaso Clinico GiardiaKarime PatiñoAún no hay calificaciones
- Medicina NuclearDocumento76 páginasMedicina NuclearDavid Jesus MoncadaAún no hay calificaciones
- Diapositiva de Espectrofotometria (Qa)Documento34 páginasDiapositiva de Espectrofotometria (Qa)Miguel Angel Huayaney Milla100% (1)
- Tecnología Médica 2Documento9 páginasTecnología Médica 2BeLeén GarciaAún no hay calificaciones
- Farmacologia Basica y Clinica Katzung 12aDocumento11 páginasFarmacologia Basica y Clinica Katzung 12aConstanza OlateAún no hay calificaciones
- 501 Tecnicas Radiologicas EspecialesDocumento8 páginas501 Tecnicas Radiologicas EspecialesWilmer AlexanderAún no hay calificaciones
- 2.2 Cuestionario. Tráfico Vesicular - ENFERMERÍADocumento3 páginas2.2 Cuestionario. Tráfico Vesicular - ENFERMERÍACielo HernandezAún no hay calificaciones
- Hayem A y Hayem BDocumento30 páginasHayem A y Hayem BJuan Domingo Davila GilAún no hay calificaciones
- Pac Desarrollo m09 RADIOFARMACIA ILERNA 9,5Documento5 páginasPac Desarrollo m09 RADIOFARMACIA ILERNA 9,5rocio.infgoAún no hay calificaciones
- RMNBNVHGVDocumento26 páginasRMNBNVHGVBrendaJazminGonzalezAún no hay calificaciones
- Informe (Toma de Muestras)Documento19 páginasInforme (Toma de Muestras)BryanLuzónAún no hay calificaciones
- Lecciones de SemiologíaDocumento143 páginasLecciones de SemiologíaGRENNY LIZANDRA CHAMBI QUISPE100% (1)
- Aritmetica en FarmacologiaDocumento41 páginasAritmetica en FarmacologiaJosselyn lissette Mancia chavezAún no hay calificaciones
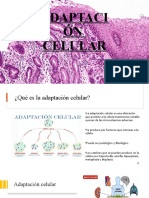
- Adaptacion Celular PresentacionDocumento26 páginasAdaptacion Celular PresentacionmilenaAún no hay calificaciones
- Sistema Inmune - Caso Clínico - HistologíaDocumento42 páginasSistema Inmune - Caso Clínico - HistologíaClaudia Cecilia Aguilar Ydiáquez100% (1)
- Elena Marin RodriguezDocumento1 páginaElena Marin RodriguezJulieta MendozaAún no hay calificaciones
- Terminología y Posicionamiento en Radiología ConvencionalDocumento48 páginasTerminología y Posicionamiento en Radiología ConvencionalRodrigo GonzalezAún no hay calificaciones
- Laboratorio 5Documento20 páginasLaboratorio 5Angie CeronAún no hay calificaciones
- Hormona PolipeptídicaDocumento2 páginasHormona PolipeptídicaMaría Isabel RicabalAún no hay calificaciones
- Enzimas en Ing. de Bioprocesos PDFDocumento145 páginasEnzimas en Ing. de Bioprocesos PDFandesonAún no hay calificaciones
- EnzimasDocumento41 páginasEnzimasreyes.sammyAún no hay calificaciones
- Tema 2.1 Enzimología ClínicaDocumento26 páginasTema 2.1 Enzimología ClínicaGuillermo LasarteAún no hay calificaciones
- 6.enzimas. Generalidades. Medida ActividadDocumento64 páginas6.enzimas. Generalidades. Medida ActividadKaren Cuenca100% (1)
- Clase de Enzimas BIO 266Documento65 páginasClase de Enzimas BIO 266KIMBERLY SAEZAún no hay calificaciones
- 6 EnzimasDocumento52 páginas6 Enzimasgianluca malerbaAún no hay calificaciones
- EncimasDocumento30 páginasEncimasRichy Q-CcAún no hay calificaciones
- ZanahoriaDocumento3 páginasZanahoriariiAún no hay calificaciones
- FRIJOLDocumento39 páginasFRIJOLriiAún no hay calificaciones
- Tomate CascaraDocumento35 páginasTomate CascarariiAún no hay calificaciones