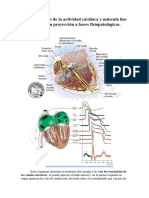
También podría gustarte
- Anatomia y Fisiologia Del Sistema Excito ConductorDocumento16 páginasAnatomia y Fisiologia Del Sistema Excito Conductorrosica1973Aún no hay calificaciones
- Mod7 TaquiarritmiasDocumento73 páginasMod7 TaquiarritmiasFernando ArancibiaAún no hay calificaciones
- ElectrofisiologíaDocumento8 páginasElectrofisiologíaAndrea Valeria Montenegro Reyes100% (1)
- CARDIO - Bases Biofísicas de La Actividad Cardiaca y Músculo Liso Vascular Con Proyección A Bases FisiopatológicasDocumento28 páginasCARDIO - Bases Biofísicas de La Actividad Cardiaca y Músculo Liso Vascular Con Proyección A Bases FisiopatológicasRomina NogueiraAún no hay calificaciones
- Génesis Del ECG NormalDocumento8 páginasGénesis Del ECG NormalCamila CarpioAún no hay calificaciones
- Electrofisiologia PDFDocumento90 páginasElectrofisiologia PDFFabiola NorabuenaAún no hay calificaciones
- Ecg CastellanosDocumento143 páginasEcg CastellanosVelia Gallegos100% (1)
- Practica Potencial de Acción CardiacoDocumento9 páginasPractica Potencial de Acción CardiacoXavi MGAún no hay calificaciones
- PRACTICA DE FISIO VIERNES - Docx CorazonnDocumento8 páginasPRACTICA DE FISIO VIERNES - Docx CorazonnlilianaAún no hay calificaciones
- Origen y Propagación 2Documento9 páginasOrigen y Propagación 2Gabriela GregórioAún no hay calificaciones
- ARRITMIASDocumento21 páginasARRITMIASJoselyn RodriguezAún no hay calificaciones
- Seminario Tp4 Fisica BiologicaDocumento8 páginasSeminario Tp4 Fisica BiologicaEstefanía MansillaAún no hay calificaciones
- Biofisica CardíacaDocumento28 páginasBiofisica CardíacaMarcos Días100% (1)
- Potencial de Membrana y CardiovasculaDocumento7 páginasPotencial de Membrana y CardiovasculaJacki ServianAún no hay calificaciones
- 07 - Propiedades Del Músculo CardiacoDocumento11 páginas07 - Propiedades Del Músculo CardiacoAlejandra HernandezAún no hay calificaciones
- 03 Guiìa Cardio ContestadaDocumento7 páginas03 Guiìa Cardio ContestadaDante ValvassoriAún no hay calificaciones
- Clase 3 4-10Documento4 páginasClase 3 4-10Valentina UbillaAún no hay calificaciones
- CARDIACODocumento67 páginasCARDIACOMayreamar EstradaAún no hay calificaciones
- Electrocardiograma Normal y Patológico - Medicina InternaDocumento35 páginasElectrocardiograma Normal y Patológico - Medicina InternaPedro HermosoAún no hay calificaciones
- UntitledDocumento7 páginasUntitledZahira AbrahamAún no hay calificaciones
- Desfibrilador CardiacoDocumento46 páginasDesfibrilador CardiacoJesus Resta NavarroAún no hay calificaciones
- Ut II Guia10Documento9 páginasUt II Guia10BarbaraAndradeQuirozAún no hay calificaciones
- BiopotencialesDocumento20 páginasBiopotencialesRoland CaixaAún no hay calificaciones
- Fisiología Cardiovascular Electrocardiografía N°4 PDFDocumento22 páginasFisiología Cardiovascular Electrocardiografía N°4 PDFKevin Argotti CazarAún no hay calificaciones
- Mecanismos Electrofisiológicos de La Elevación Del Segmento ST Durante El Infarto Agudo de Miocardio. Hipótesis ActualDocumento7 páginasMecanismos Electrofisiológicos de La Elevación Del Segmento ST Durante El Infarto Agudo de Miocardio. Hipótesis ActualElíseo E LQAún no hay calificaciones
- Tarea 2 - Miguel Angel Ahumada Segura!!!Documento16 páginasTarea 2 - Miguel Angel Ahumada Segura!!!MIGUEL ANGEL AHUMADAAún no hay calificaciones
- Manual Toma e Interpretacion de ElectrocardiogramaDocumento29 páginasManual Toma e Interpretacion de ElectrocardiogramaGuillermina SanchezAún no hay calificaciones
- Aparato Cardiovascular 1T1Documento41 páginasAparato Cardiovascular 1T1Marycielo LmAún no hay calificaciones
- Fisiología - Guía # 1. Actividad Electrica Cardiaca 2023-2Documento10 páginasFisiología - Guía # 1. Actividad Electrica Cardiaca 2023-2LAURA SOFIA PEREZ CUELLOAún no hay calificaciones
- CARDIO!Documento31 páginasCARDIO!Mauricio VianaAún no hay calificaciones
- Propiedades Del Corazon de Sapo - CompressDocumento23 páginasPropiedades Del Corazon de Sapo - CompressJUAN RELUZ ZAMBRANOAún no hay calificaciones
- Electrofisiología CardíacaDocumento13 páginasElectrofisiología CardíacajohanaAún no hay calificaciones
- 1 EkgDocumento12 páginas1 EkgegleeAún no hay calificaciones
- Teoria Delm Musculo CardiacoDocumento12 páginasTeoria Delm Musculo CardiacoAlbert KantAún no hay calificaciones
- Marco Teorico ECGDocumento11 páginasMarco Teorico ECGEliana Caceres TorricoAún no hay calificaciones
- EKG Normal 1 y 2Documento12 páginasEKG Normal 1 y 2Franck FarreraAún no hay calificaciones
- GUÍA DE PRACTICA 3 Efecto de Los Iones Sobre La Frecuencia CardíacaDocumento4 páginasGUÍA DE PRACTICA 3 Efecto de Los Iones Sobre La Frecuencia CardíacaArely Mical SánchezAún no hay calificaciones
- Documento Sin TítuloDocumento15 páginasDocumento Sin TítuloGuillermina SanchezAún no hay calificaciones
- MielinizacionDocumento57 páginasMielinizacionVictoria Castillo100% (1)
- Electrofisiologia CardiacaDocumento14 páginasElectrofisiologia Cardiacaluispin1186Aún no hay calificaciones
- Electrofisiología y EcgDocumento11 páginasElectrofisiología y Ecgivanlalvarez.22Aún no hay calificaciones
- Automatismo AnormalDocumento3 páginasAutomatismo AnormalEmily Janice Oliva AlburquequeAún no hay calificaciones
- Propiedades Electricas Del CorazonDocumento4 páginasPropiedades Electricas Del CorazonRichard Armijo GarridoAún no hay calificaciones
- Secme-4237 1Documento11 páginasSecme-4237 1JoséAún no hay calificaciones
- Electrofisiologia Cardiaca CMDM PDFDocumento10 páginasElectrofisiologia Cardiaca CMDM PDFWendy OrtizAún no hay calificaciones
- Biofísica Cardiaca - YGDocumento3 páginasBiofísica Cardiaca - YGmichaellaAún no hay calificaciones
- BiopotencialesDocumento24 páginasBiopotencialesMiguel SoberanesAún no hay calificaciones
- Tarea 2 - Trabajo ColaborativoDocumento46 páginasTarea 2 - Trabajo ColaborativoJuan Carlos Grisales MenesesAún no hay calificaciones
- Electrocardiograma IDocumento19 páginasElectrocardiograma IGeraldine LópezAún no hay calificaciones
- UTII 1 Practica Potencial de Accion CardiacoDocumento13 páginasUTII 1 Practica Potencial de Accion CardiacoMarisol RodríguezAún no hay calificaciones
- Cardiología CTODocumento95 páginasCardiología CTOEfren Penaflores100% (1)
- Apunte EelctrocardiografiaDocumento84 páginasApunte EelctrocardiografiaMelissa JerezAún no hay calificaciones
- Teoría-Función Cardiovascular - MEHU 632Documento24 páginasTeoría-Función Cardiovascular - MEHU 632Sara ParedesAún no hay calificaciones
- Electrocardiograma 01Documento15 páginasElectrocardiograma 01Yasirys Mariell PichardoAún no hay calificaciones
- Cómo se comunican las neuronas: El milagro de la transmisión sinápticaDe EverandCómo se comunican las neuronas: El milagro de la transmisión sinápticaAún no hay calificaciones
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- Medicina energética: Acupuntura 3 Circuitos energéticos secundariosDe EverandMedicina energética: Acupuntura 3 Circuitos energéticos secundariosCalificación: 5 de 5 estrellas5/5 (5)
- Caja - Corazón 5Documento17 páginasCaja - Corazón 5DeniseDelPuertoAún no hay calificaciones
- Arritmias Fajuri 2009Documento15 páginasArritmias Fajuri 2009jorge51988Aún no hay calificaciones
- Ciclo Eléctrico Del Corazón1Documento5 páginasCiclo Eléctrico Del Corazón1Dalia Gomez GarciaAún no hay calificaciones
- El CorazónDocumento3 páginasEl CorazónCarmen RojasAún no hay calificaciones
- Problemas de BioelectricidadDocumento9 páginasProblemas de BioelectricidadSofiaAún no hay calificaciones
- Sistema de Estimulación y Conducción Del CorazónDocumento11 páginasSistema de Estimulación y Conducción Del CorazónGabyta BarreraAún no hay calificaciones
- Bases Fisiologicas de Las CardiopatiasDocumento480 páginasBases Fisiologicas de Las CardiopatiasLeidy Nayerli Garcia Rodriguez100% (1)
- CFC - Corazón 5Documento18 páginasCFC - Corazón 5Karen RodríguezAún no hay calificaciones
- Sistema Electrico Del CorazónDocumento3 páginasSistema Electrico Del CorazónFlor MartinezAún no hay calificaciones
- Sem 4. Presentación1 ElectroDocumento16 páginasSem 4. Presentación1 ElectrolauracAún no hay calificaciones
- Sistema de Conducción CardiacoDocumento3 páginasSistema de Conducción CardiacoVicente Jr Padilla0% (1)
- Cardiologia Bases Fisiopatologicas de Las CardiopatiasDocumento478 páginasCardiologia Bases Fisiopatologicas de Las CardiopatiasismaelcosioAún no hay calificaciones
- Resumen de Corazon RouviereDocumento35 páginasResumen de Corazon RouviereRamon Martunez LoraAún no hay calificaciones
- Ligaduras de StaDocumento11 páginasLigaduras de StaSELENE VANESSA CHOZO MESTANZAAún no hay calificaciones
- Anatomia 05 CardiovascularDocumento52 páginasAnatomia 05 CardiovascularAlexis SccAún no hay calificaciones
- UACV - Sistema de ConduccionDocumento7 páginasUACV - Sistema de ConduccionsotelofrancoezequielAún no hay calificaciones
- Descripción Del Sistema de Exitoconducción CardiacaDocumento2 páginasDescripción Del Sistema de Exitoconducción CardiacaYuvixa NavaAún no hay calificaciones
- MorfofisiologiaDocumento13 páginasMorfofisiologia¡Hey Soy Patto100% (2)
- Fibras de PurkinjeDocumento5 páginasFibras de PurkinjeLishii Ðë Lä CallëAún no hay calificaciones
- Vias IntranodularesDocumento7 páginasVias IntranodularesnatalyAún no hay calificaciones
- Electrofisiologia 1 PDFDocumento12 páginasElectrofisiologia 1 PDFAlejandro Pari VillenaAún no hay calificaciones
- Como Esta Formado El Aparato CardiovascularDocumento10 páginasComo Esta Formado El Aparato CardiovascularDisami Salazar MioAún no hay calificaciones
- ANATOMIA HUMANA RecensiónDocumento4 páginasANATOMIA HUMANA Recensiónjuan bautista acosta aquinoAún no hay calificaciones
- Ciclo CardiacoDocumento56 páginasCiclo CardiacoCamilo HenaoAún no hay calificaciones
- Sistema Nodal y Conducción PDFDocumento18 páginasSistema Nodal y Conducción PDFMaría Yanet Rodrìguez Soberòn100% (1)
- Sistema Excito-ConductorDocumento3 páginasSistema Excito-ConductorMartín Torreblanca RodriguezAún no hay calificaciones
- FARMACOLOGIA Digitalicos y AntiarritmicosDocumento55 páginasFARMACOLOGIA Digitalicos y AntiarritmicosCleysser Antonio Custodio Polar100% (1)
- Cap III Cardio 2Documento17 páginasCap III Cardio 2Anonymous alzR2b1pqAún no hay calificaciones