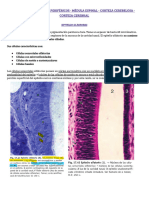
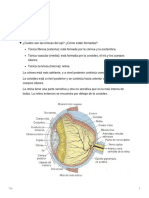
También podría gustarte
- Presentación Anatomia y Fisiologia OjoDocumento27 páginasPresentación Anatomia y Fisiologia OjoVictor Winchester100% (1)
- Manual Uñas AcrilicasDocumento83 páginasManual Uñas AcrilicasFinarep PerezAún no hay calificaciones
- Fisiologia de Los Sentidos CompletoDocumento17 páginasFisiologia de Los Sentidos CompletoJeavAún no hay calificaciones
- El Ojo Humano Origen Desarrollo Vision Monocular 6850 CompletoDocumento29 páginasEl Ojo Humano Origen Desarrollo Vision Monocular 6850 CompletoMarion FernandaAún no hay calificaciones
- Histología y Embrología del Sistema NerviosoDe EverandHistología y Embrología del Sistema NerviosoCalificación: 3 de 5 estrellas3/5 (1)
- Organos de Los SentidosDocumento16 páginasOrganos de Los SentidosMaria Luisa Paucar RomeroAún no hay calificaciones
- Órganos de Los SentidosDocumento9 páginasÓrganos de Los SentidosFacsury Sanjuan MezaAún no hay calificaciones
- Cristian Gonzalez Semana 9 Proyecto FinalDocumento10 páginasCristian Gonzalez Semana 9 Proyecto FinalCristian GonzálezAún no hay calificaciones
- Órgano de La Visión AnatomíaDocumento12 páginasÓrgano de La Visión AnatomíanikolettsmirnofffaAún no hay calificaciones
- Sistema de Los SentidosDocumento15 páginasSistema de Los SentidosRaul GonzalezAún no hay calificaciones
- Histo SentidosDocumento7 páginasHisto Sentidoshugo rojasAún no hay calificaciones
- Sistemas Sensoriales Gusto y OlfatoDocumento7 páginasSistemas Sensoriales Gusto y OlfatoisabellaAún no hay calificaciones
- Percepcion SensorialDocumento14 páginasPercepcion SensorialShamy ShamyAún no hay calificaciones
- El Ojo HumanoDocumento18 páginasEl Ojo HumanoJose Manuel Blazquez BlancoAún no hay calificaciones
- Actividad 3 MorfoDocumento8 páginasActividad 3 MorfoKaren PeñalozaAún no hay calificaciones
- Histología Ii. Mc. Rse.Documento12 páginasHistología Ii. Mc. Rse.Amarildo CristinoAún no hay calificaciones
- Conos y BastonesDocumento2 páginasConos y BastonesDANIEL TOVARAún no hay calificaciones
- Resumen Módulo II - 2023 Receptores SensorialesDocumento17 páginasResumen Módulo II - 2023 Receptores Sensorialesmartin.urri2898Aún no hay calificaciones
- Texto VisionDocumento2 páginasTexto VisionTía Marité LagosAún no hay calificaciones
- El Ojo Comienza A Formarse A Partir Del Desarrollo EmbrionarioDocumento12 páginasEl Ojo Comienza A Formarse A Partir Del Desarrollo EmbrionarioMarisa Niticia Noya CalviñoAún no hay calificaciones
- Capitulo IiDocumento19 páginasCapitulo Iijuan achaAún no hay calificaciones
- Sentido de La Vista en AnimalesDocumento7 páginasSentido de La Vista en AnimalesLi IzzAún no hay calificaciones
- OJODocumento14 páginasOJOAlejandro BedollaAún no hay calificaciones
- A Retina Es El Órgano Sensorial Del OjoDocumento4 páginasA Retina Es El Órgano Sensorial Del OjoLuisa FERNANDAAún no hay calificaciones
- OjoDocumento17 páginasOjoGonzalo ManavellaAún no hay calificaciones
- COROIDESDocumento3 páginasCOROIDESLeary UchethAún no hay calificaciones
- Resumen Fisiologia de La VisionDocumento2 páginasResumen Fisiologia de La VisionAndres ChachaAún no hay calificaciones
- Anatomia Ocular Leccion 5Documento46 páginasAnatomia Ocular Leccion 5emilyzavala1143009Aún no hay calificaciones
- Bloque 2 AnatomiaDocumento83 páginasBloque 2 AnatomiaRuben ZamoraAún no hay calificaciones
- Anat Aplic Los Sentidos 1º Bach Univ LabDocumento15 páginasAnat Aplic Los Sentidos 1º Bach Univ Labjuancarloscamposromero173Aún no hay calificaciones
- Aparato de La VisionDocumento3 páginasAparato de La Visionraul escobarAún no hay calificaciones
- Biologia 3actDocumento4 páginasBiologia 3actRosario HernandezAún no hay calificaciones
- Histología Del OjoDocumento24 páginasHistología Del OjoLucia Saavedra LozadaAún no hay calificaciones
- Word REtinaDocumento89 páginasWord REtinaCesar Alex Lumbre YuptonAún no hay calificaciones
- S. 1 C.A IiDocumento33 páginasS. 1 C.A IiNachooAún no hay calificaciones
- S. 1 C.A Ii-1-9Documento9 páginasS. 1 C.A Ii-1-9NachooAún no hay calificaciones
- EstesiologiaDocumento22 páginasEstesiologiaYurani RojasAún no hay calificaciones
- Clase 1 MFDocumento6 páginasClase 1 MFConichpAún no hay calificaciones
- Tarea 3 - Via Visual PDFDocumento30 páginasTarea 3 - Via Visual PDFJaimeSergioVasquezZunigaAún no hay calificaciones
- Visión - Wikipedia, La Enciclopedia LibreDocumento9 páginasVisión - Wikipedia, La Enciclopedia LibreBeto GomitoAún no hay calificaciones
- Diapositivas de Anatomia Ocular 2013Documento20 páginasDiapositivas de Anatomia Ocular 2013Lourdes Gallego VilcherrezAún no hay calificaciones
- Generalidades Del Globo OcularDocumento4 páginasGeneralidades Del Globo OcularÁngelaZemamaAún no hay calificaciones
- Organo de La VisionDocumento33 páginasOrgano de La Visionjunior pinedaAún no hay calificaciones
- Resumen de OjoDocumento3 páginasResumen de OjoLuisFelipe Saldías0% (2)
- Sem 16-17 Fisiología y Envejecimiento de Los SentidosDocumento59 páginasSem 16-17 Fisiología y Envejecimiento de Los Sentidos05-EF-HU-PATRICIA ARTEAGA ORDOÑEZAún no hay calificaciones
- Informe Cienticfico Ojo VacunoDocumento15 páginasInforme Cienticfico Ojo VacunoCarlos Felipe Rojas ParraAún no hay calificaciones
- BBIOLOGGIA 3 MedioDocumento4 páginasBBIOLOGGIA 3 Mediogaby cabreraAún no hay calificaciones
- El OjoDocumento9 páginasEl OjoBeatrizAún no hay calificaciones
- El Ojo Es Un Órgano Sensorial Complejo Que Actúa Como Receptor Del Aparato de La VisiónDocumento5 páginasEl Ojo Es Un Órgano Sensorial Complejo Que Actúa Como Receptor Del Aparato de La VisiónYasna Isabel Molina GuajardoAún no hay calificaciones
- 4º Ficha - Sistema-Sensorial 04-07-23 RESUELTODocumento9 páginas4º Ficha - Sistema-Sensorial 04-07-23 RESUELTOLiz AquijeAún no hay calificaciones
- 4º Ficha - Sistema-Sensorial 04-07-23Documento8 páginas4º Ficha - Sistema-Sensorial 04-07-23Aparcana MedinaAún no hay calificaciones
- Primer Taller de OftalmologíaDocumento8 páginasPrimer Taller de Oftalmologíamedicinafusm aybAún no hay calificaciones
- Histologia de Ojo - RetinaDocumento19 páginasHistologia de Ojo - RetinadalianajaelgonzalezAún no hay calificaciones
- Psicofisiologia Repaso 2Documento9 páginasPsicofisiologia Repaso 2camilamartinez2572004Aún no hay calificaciones
- Anatomia y Fisiologia de Los Órganos de PDFDocumento44 páginasAnatomia y Fisiologia de Los Órganos de PDFmaria luzvenita vittery gonzalesAún no hay calificaciones
- Organos de Los Sentidos (Ojo)Documento13 páginasOrganos de Los Sentidos (Ojo)Abraham GuidoAún no hay calificaciones
- Los Órganos de Los SentidosDocumento54 páginasLos Órganos de Los SentidoshelenemeyersAún no hay calificaciones
- Taller de Histologia. Sentidos Especiales AdrianaDocumento18 páginasTaller de Histologia. Sentidos Especiales AdrianaJuan David AlarcónAún no hay calificaciones
- Ojo HumanoDocumento12 páginasOjo HumanoIngrid Tatiana HolguinAún no hay calificaciones
- Fundamentos Físicos de La VisiónDocumento14 páginasFundamentos Físicos de La VisiónLaura RamirezAún no hay calificaciones
- Tema 2 Psicologia FisiologicaDocumento11 páginasTema 2 Psicologia FisiologicaAlvaro Garrido GarciaAún no hay calificaciones
- ÓRGANOS DE LOS SENTIDOS ApuntesDocumento9 páginasÓRGANOS DE LOS SENTIDOS ApuntesJuan Manuel GuerreroAún no hay calificaciones
- Manejo de RpbiDocumento13 páginasManejo de RpbiAurora Parissi VicenteAún no hay calificaciones
- IBM II 2020 2.3.2 G14 Ejercicico Clínico Patológico - GJMDocumento4 páginasIBM II 2020 2.3.2 G14 Ejercicico Clínico Patológico - GJMGuillermo JuarezAún no hay calificaciones
- Farmacocinética 2.0Documento6 páginasFarmacocinética 2.0Guillermo JuarezAún no hay calificaciones
- Duelo Normal y Patológico en Los Familiares y AmigosDocumento4 páginasDuelo Normal y Patológico en Los Familiares y AmigosGuillermo JuarezAún no hay calificaciones
- Cáncer PulmonarDocumento18 páginasCáncer PulmonarGuillermo JuarezAún no hay calificaciones
- 2.2.2 Salud Sexual y Reproductiva en Los Adolescentes - MFMBDocumento4 páginas2.2.2 Salud Sexual y Reproductiva en Los Adolescentes - MFMBGuillermo JuarezAún no hay calificaciones
- 2.3.1 Salud y Enfermedad Durante La Vida Adulta - MFMBDocumento5 páginas2.3.1 Salud y Enfermedad Durante La Vida Adulta - MFMBGuillermo JuarezAún no hay calificaciones
- 2.2.1 Salud Sexual y Reproductiva en Los AdolescentesDocumento4 páginas2.2.1 Salud Sexual y Reproductiva en Los AdolescentesGuillermo JuarezAún no hay calificaciones
- 2.3.1 Salud y Enfermedad Durante La Vida Adulta - MFMBDocumento5 páginas2.3.1 Salud y Enfermedad Durante La Vida Adulta - MFMBGuillermo JuarezAún no hay calificaciones
- 2.2.2 Salud Sexual y Reproductiva en Los Adolescentes - MFMBDocumento4 páginas2.2.2 Salud Sexual y Reproductiva en Los Adolescentes - MFMBGuillermo JuarezAún no hay calificaciones
- 1.2 PrácticaAnatomíaDocumento8 páginas1.2 PrácticaAnatomíaGuillermo JuarezAún no hay calificaciones
- Trastorno AntisocialDocumento5 páginasTrastorno AntisocialGuillermo JuarezAún no hay calificaciones
- Itv-Vi-Po-002-07 Rev. 1Documento1 páginaItv-Vi-Po-002-07 Rev. 1Guillermo JuarezAún no hay calificaciones
- Sesión 01 TFDocumento79 páginasSesión 01 TFNícolas LeyvaAún no hay calificaciones
- El Trabajo en El LaboratorioDocumento3 páginasEl Trabajo en El LaboratorioMaria Luengo LahozAún no hay calificaciones
- Lütze Superflex Tronic (C) Pur - Ul - 4xu04-50228 (00000003)Documento3 páginasLütze Superflex Tronic (C) Pur - Ul - 4xu04-50228 (00000003)Marco ApellidoAún no hay calificaciones
- Desinfeccion y Limpieza en El LaboratorioDocumento1 páginaDesinfeccion y Limpieza en El LaboratorioX17 1Aún no hay calificaciones
- MSDS Once PinturaDocumento8 páginasMSDS Once PinturaCeleste Espejo VillegasAún no hay calificaciones
- Tesis Propuesta Metodologica Basada en La Innovacion para La Ensenanza-Aprendizaje de La Unidad Teoria Del Enlace en La - 930 - 651u.image - MarkedDocumento85 páginasTesis Propuesta Metodologica Basada en La Innovacion para La Ensenanza-Aprendizaje de La Unidad Teoria Del Enlace en La - 930 - 651u.image - MarkedMartina Antonella MadridAún no hay calificaciones
- Reporte 6. Reaccion en Estado SolidoDocumento6 páginasReporte 6. Reaccion en Estado SolidoALDO DANIEL GONZALEZ RESENDIZAún no hay calificaciones
- Diagrama Sipoc y Fichas de ProcesosDocumento10 páginasDiagrama Sipoc y Fichas de ProcesosDarkness jhlpAún no hay calificaciones
- Capitulo Dos: Inspección de Soldaduras IiDocumento40 páginasCapitulo Dos: Inspección de Soldaduras IiHenry GonzalezAún no hay calificaciones
- Informe de Laboratorio PHDocumento2 páginasInforme de Laboratorio PHcamila moncayoAún no hay calificaciones
- Informe Practica 2Documento4 páginasInforme Practica 2CRISTIAN DAVID ARIZA OROZCO100% (1)
- Biofisica Respiratoria 2022Documento23 páginasBiofisica Respiratoria 2022Estrella GonzalezAún no hay calificaciones
- FTTH - Especificaciones Técnicas de Las Orejas de Anclaje Con Ganchos De...Documento2 páginasFTTH - Especificaciones Técnicas de Las Orejas de Anclaje Con Ganchos De...Jean Franco Calderon SosaAún no hay calificaciones
- Clase 1-FarmaciaDocumento37 páginasClase 1-FarmaciaPercy Paucar CuevaAún no hay calificaciones
- Acidez de La Leche Marco TeoricoDocumento3 páginasAcidez de La Leche Marco TeoricoJoseline EblaAún no hay calificaciones
- Marca Pe Cemento Material Pe Agregado TMN MF Wa Wo Varillado Suelto Suelto Arena Piedra Propiedades Peso Unitario SecoDocumento5 páginasMarca Pe Cemento Material Pe Agregado TMN MF Wa Wo Varillado Suelto Suelto Arena Piedra Propiedades Peso Unitario SecoAlessandra GlenniAún no hay calificaciones
- El PH y Su ImportanciaDocumento3 páginasEl PH y Su ImportanciaJOHAN ABELARDO JESUS CASTRO ESTRADA0% (1)
- Factores Que Afectan El Crecimiento MicrobianoDocumento4 páginasFactores Que Afectan El Crecimiento MicrobianoMÓNICA MARÍN ARBELÁEZAún no hay calificaciones
- Analisis, Interp. Recom. de Suelos Yonathan GarciaDocumento6 páginasAnalisis, Interp. Recom. de Suelos Yonathan GarciaValentina AmarilesAún no hay calificaciones
- Dureza Del AguaDocumento14 páginasDureza Del AguaRodrigo RicoAún no hay calificaciones
- TD-CON-PETS-011 Preparacion ReactivosDocumento4 páginasTD-CON-PETS-011 Preparacion ReactivoscarlosAún no hay calificaciones
- Ejercicio EjesDocumento17 páginasEjercicio EjesIver SanchezAún no hay calificaciones
- Ilovepdf Merged 2Documento18 páginasIlovepdf Merged 2Pao UrrutiaAún no hay calificaciones
- Suspensión (Química) - Wikipedia, La Enciclopedia LibreDocumento3 páginasSuspensión (Química) - Wikipedia, La Enciclopedia Libregearfield07Aún no hay calificaciones
- Tarea 4 (Formulaciòn y Nomenclatura de Compuestos)Documento6 páginasTarea 4 (Formulaciòn y Nomenclatura de Compuestos)Mauricio Araujo AriasAún no hay calificaciones
- Blue PointDocumento6 páginasBlue PointGustavo Vallejos AdrianzénAún no hay calificaciones
- Fiq Sa ReyesDocumento103 páginasFiq Sa ReyesDaniel Torres TorresAún no hay calificaciones
- Informe ABSORCION PDFDocumento21 páginasInforme ABSORCION PDFDavid Camilo Guerrero GonzalezAún no hay calificaciones