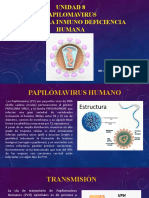
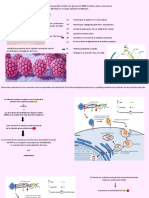
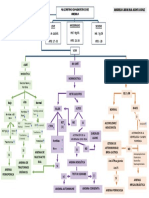
También podría gustarte
- Biologiadelvph 150914030928 Lva1 App6892Documento54 páginasBiologiadelvph 150914030928 Lva1 App6892Coral Garcia RiveraAún no hay calificaciones
- Virus del Papiloma Humano (HPV) Causas, Tipos y Factores de RiesgoDocumento14 páginasVirus del Papiloma Humano (HPV) Causas, Tipos y Factores de RiesgoMireya VelascoAún no hay calificaciones
- Oportunidades de Diagnóstico Molecular para El Cáncer CervicouterinoDocumento30 páginasOportunidades de Diagnóstico Molecular para El Cáncer CervicouterinoDaniel Alejandro Estrada MondragónAún no hay calificaciones
- Virus Del Papiloma Humano (VPH)Documento44 páginasVirus Del Papiloma Humano (VPH)Eduardo Gómez100% (1)
- VirusDocumento19 páginasVirusNoelia LeonAún no hay calificaciones
- Fisiopatologia Infeccion Por VPHDocumento29 páginasFisiopatologia Infeccion Por VPHClaudia Mercedes Alonso Cuevas50% (2)
- Monografia VPH.Documento5 páginasMonografia VPH.Kevin SánchezAún no hay calificaciones
- Virus Del Papiloma Humano VPH Cito3erparcialDocumento5 páginasVirus Del Papiloma Humano VPH Cito3erparcialRosario Calani CanaviriAún no hay calificaciones
- Papovavirus: Familia PapovaviridaeDocumento3 páginasPapovavirus: Familia PapovaviridaeAnonymous TN4XtXvAún no hay calificaciones
- Biologiadelvph 150914030928 Lva1 App6892Documento54 páginasBiologiadelvph 150914030928 Lva1 App6892Jersinho medina llerenaAún no hay calificaciones
- Papiloma Humano Trabajo OficialDocumento8 páginasPapiloma Humano Trabajo OficialVicenteRamosMamaniAún no hay calificaciones
- Virus Tumorales y OncogenesDocumento11 páginasVirus Tumorales y OncogenesJhoann Sebastian CucaitaAún no hay calificaciones
- 4-Interacción Virus CelulaDocumento4 páginas4-Interacción Virus CelulaGeronimo TognottiAún no hay calificaciones
- ReplicaciónDocumento3 páginasReplicaciónSebastian ManriqueAún no hay calificaciones
- Antiherpeticos guión expo 2Documento34 páginasAntiherpeticos guión expo 2camilalopcepAún no hay calificaciones
- Retrovirus (VIH) Principios de Microbiologia Murray 7Documento8 páginasRetrovirus (VIH) Principios de Microbiologia Murray 7Daniel GarciaAún no hay calificaciones
- 431-Texto Del Artículo-769-2-10-20210407Documento17 páginas431-Texto Del Artículo-769-2-10-20210407Brenda Hernández TrejoAún no hay calificaciones
- CUESTIONARIO VPH - Guadalupe Herrera RamosDocumento3 páginasCUESTIONARIO VPH - Guadalupe Herrera RamosGuadalupe Herrera RamosAún no hay calificaciones
- Virus Del Papiloma HumanoDocumento32 páginasVirus Del Papiloma HumanoCesar EspinozaAún no hay calificaciones
- Virus Del Papiloma Humano y Cancer de Cuello Uterino.Documento27 páginasVirus Del Papiloma Humano y Cancer de Cuello Uterino.Rbk JGAún no hay calificaciones
- HerpesvirusDocumento81 páginasHerpesvirusegmar06Aún no hay calificaciones
- Unidad 8 Papilomavirus - VihDocumento18 páginasUnidad 8 Papilomavirus - VihLISBETH PANIAGUA GUTIERREZAún no hay calificaciones
- TareahepatitisDocumento3 páginasTareahepatitisAlexandra Stephanie Santa Cruz BellidoAún no hay calificaciones
- VPHDocumento21 páginasVPHYessmi Ramos Miranda100% (2)
- Virus Papiloma Humano (VPH)Documento10 páginasVirus Papiloma Humano (VPH)CLaudia Julieta Robles PLanellsAún no hay calificaciones
- Cap 8 Parte 4Documento12 páginasCap 8 Parte 4Nini Sierra GutierrezAún no hay calificaciones
- Teo 7 ImprimirDocumento3 páginasTeo 7 ImprimirConstanza Mariel AmerioAún no hay calificaciones
- Marco Teorico PreDocumento16 páginasMarco Teorico PreMiguel VGAún no hay calificaciones
- Presentación1 (Autoguardado)Documento5 páginasPresentación1 (Autoguardado)Monica MartinezAún no hay calificaciones
- Imagenes en MedicinaDocumento2 páginasImagenes en MedicinaLucía Maldonado De LeónAún no hay calificaciones
- HerpesDocumento25 páginasHerpesPatricia MontemiraAún no hay calificaciones
- VPHDocumento2 páginasVPHSOFIA ALEJANDRA SANABRIA FANDIÑOAún no hay calificaciones
- Introducción A La VirologíaDocumento15 páginasIntroducción A La VirologíaSietecolores ReyAún no hay calificaciones
- Virología Del VIH PDFDocumento32 páginasVirología Del VIH PDFYamil ReyesAún no hay calificaciones
- Virologia: Universidad de Guayaquil Facultad de Ciencias Médicas Carrera de MedicinaDocumento28 páginasVirologia: Universidad de Guayaquil Facultad de Ciencias Médicas Carrera de MedicinaBigi MercuryAún no hay calificaciones
- VIH, Resumen FinalDocumento10 páginasVIH, Resumen FinalEduardo MiramontesAún no hay calificaciones
- 2 PapillomavirusDocumento48 páginas2 PapillomavirusPriscila ElizabethAún no hay calificaciones
- VIHDocumento19 páginasVIHandres barretoAún no hay calificaciones
- Estructura y epidemiología del VIHDocumento15 páginasEstructura y epidemiología del VIHPolSejLop LópezAún no hay calificaciones
- Virus de Papiloma Humano SEMIOLOGIADocumento3 páginasVirus de Papiloma Humano SEMIOLOGIAAzul ChinteteAún no hay calificaciones
- Material de Estudio - VIH-SIDADocumento19 páginasMaterial de Estudio - VIH-SIDAManecitaGarcíaAún no hay calificaciones
- Mod II-01 Texto Virologia 2021-2022Documento33 páginasMod II-01 Texto Virologia 2021-2022Stephanie Orozco CastilloAún no hay calificaciones
- Virus Del Papiloma HumanoDocumento5 páginasVirus Del Papiloma HumanoSofia RamosAún no hay calificaciones
- RetrovirusDocumento9 páginasRetrovirusfranchescaramirezAún no hay calificaciones
- VIH 2016 MicroDocumento24 páginasVIH 2016 MicroRicardo AguileraAún no hay calificaciones
- Herpesvirus: estructura, ciclo de vida y manifestaciones clínicasDocumento13 páginasHerpesvirus: estructura, ciclo de vida y manifestaciones clínicasClay MukurnereAún no hay calificaciones
- Anestesia e Infección Por HIVDocumento27 páginasAnestesia e Infección Por HIVmalusetti100% (20)
- Extension de Bio-mol 1 Er Parcial 2024Documento13 páginasExtension de Bio-mol 1 Er Parcial 2024Gaby Rojas pardoAún no hay calificaciones
- Infecciones persistentesDocumento16 páginasInfecciones persistentesMelina BritezAún no hay calificaciones
- Virus CompletoDocumento21 páginasVirus CompletoEthel CoriaAún no hay calificaciones
- Vacunas contra el VPH: Características y mecanismo de acciónDocumento26 páginasVacunas contra el VPH: Características y mecanismo de acciónDemyMichMoncada100% (1)
- Herpes SeleDocumento27 páginasHerpes SeleSelene Vanessa Chozo MestanzaAún no hay calificaciones
- Patogenia Del VPHDocumento2 páginasPatogenia Del VPHAlexandra perez gamonalAún no hay calificaciones
- Resumen Cap. 51 MicrobiologiaDocumento5 páginasResumen Cap. 51 MicrobiologiaMadelyn Leal del ValleAún no hay calificaciones
- File 2Documento11 páginasFile 2Alexis FernandoAún no hay calificaciones
- 3 virus pielDocumento4 páginas3 virus pielYeisy Ruiz SierraAún no hay calificaciones
- Teoria N°9 Respuestas IiiDocumento51 páginasTeoria N°9 Respuestas IiiANAAún no hay calificaciones
- Síndrome de Wiskott Aldrich: causas, síntomas y herenciaDocumento12 páginasSíndrome de Wiskott Aldrich: causas, síntomas y herenciavalentina medina hernandezAún no hay calificaciones
- Bioquímica de La DiabetesDocumento20 páginasBioquímica de La DiabetesIsaac Palacios50% (2)
- Tabla de MetaloenzimasDocumento2 páginasTabla de MetaloenzimasJose RSAún no hay calificaciones
- Clase QPCR 2020 Daniela CapiatiDocumento40 páginasClase QPCR 2020 Daniela CapiatiNatalia Andrea Nuñez ToledoAún no hay calificaciones
- El Ciclo CelularDocumento6 páginasEl Ciclo Celulargabriela muñoz100% (1)
- Ejercicios de Transporte A Travez de La Membrana CelularDocumento5 páginasEjercicios de Transporte A Travez de La Membrana CelularCarlos SantiagoAún no hay calificaciones
- Pared CelularDocumento9 páginasPared CelularmieleritoAún no hay calificaciones
- Fisiología del shock insulínicoDocumento27 páginasFisiología del shock insulínicoBenito Rosas100% (2)
- Identificación Cualitativa de CarbohidratosDocumento4 páginasIdentificación Cualitativa de CarbohidratosJorleny Jamileth Marquez CamachoAún no hay calificaciones
- CitigenéticaDocumento3 páginasCitigenéticaCarlos AguillónAún no hay calificaciones
- Radicales Libres y Su Incidencia en El EnvejecimientoDocumento15 páginasRadicales Libres y Su Incidencia en El EnvejecimientoMario FierroAún no hay calificaciones
- Regulacion EnzimaticaDocumento5 páginasRegulacion EnzimaticaCitlalic Lizeth GavilanesAún no hay calificaciones
- Sistema Nervioso 1Documento41 páginasSistema Nervioso 1valeAún no hay calificaciones
- Estructura Quimica Del AdnDocumento9 páginasEstructura Quimica Del AdnPedro Roger LucesAún no hay calificaciones
- Actividades Clase 5 Enzimas Parte 2 Unidad 1Documento2 páginasActividades Clase 5 Enzimas Parte 2 Unidad 1Ingrid YánezAún no hay calificaciones
- TEMA 11 Transcripción y TraducciónDocumento8 páginasTEMA 11 Transcripción y TraducciónLAURA LARAAún no hay calificaciones
- Algoritmo Anemia TerminadoDocumento1 páginaAlgoritmo Anemia TerminadoANDREA CAROLINA ACOSTA CRUZAún no hay calificaciones
- Mesoterapia Facial 222Documento14 páginasMesoterapia Facial 222Dagmarys100% (3)
- Mapa Mental (Neurotrasmisor Acetilcolina) .Documento4 páginasMapa Mental (Neurotrasmisor Acetilcolina) .Andres eduardo Sanchez nossaAún no hay calificaciones
- I. Seminarios Cinética EnzimáticaDocumento7 páginasI. Seminarios Cinética EnzimáticaLaura Célix100% (1)
- Informe Elaboracion de Un Adn de Papel-G-B1Documento14 páginasInforme Elaboracion de Un Adn de Papel-G-B1miriam arocutipa ticonaAún no hay calificaciones
- La Tre 4Documento14 páginasLa Tre 4JACKELINE ROXANA HUAICHO AMARAún no hay calificaciones
- Recuperación Sexto y Septimo 2023Documento1 páginaRecuperación Sexto y Septimo 2023pajarita45Aún no hay calificaciones
- Citoesqueleto Microtúbulos PDFDocumento7 páginasCitoesqueleto Microtúbulos PDFCesar AvalosAún no hay calificaciones
- Cuestionario 4. Las Mitocondrias y La Respiración AeróbicaDocumento4 páginasCuestionario 4. Las Mitocondrias y La Respiración AeróbicaIncógnito MedAún no hay calificaciones
- Preinforme EnzimasDocumento15 páginasPreinforme EnzimasDavid RodriguezAún no hay calificaciones
- Cuadro de Planificacion 2023-2024. Biologia de 5toDocumento2 páginasCuadro de Planificacion 2023-2024. Biologia de 5toGAUDDY0909Aún no hay calificaciones
- Taller Individual La CélulaDocumento2 páginasTaller Individual La CélulaclaudiaAún no hay calificaciones
- Silabo Bioquimica y Nutricion 2020-I - 20200311084924Documento21 páginasSilabo Bioquimica y Nutricion 2020-I - 20200311084924Alex OrozcoAún no hay calificaciones
- Introducción a la bioquímica de alimentos: lípidosDocumento38 páginasIntroducción a la bioquímica de alimentos: lípidosLuis Eduardo Gomez GuevaraAún no hay calificaciones
- Comprension Lectora La RespiracionDocumento2 páginasComprension Lectora La RespiracionJazmin Aguirre HenaoAún no hay calificaciones