También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Clase 4 Unidad 3 - ReplicaciónDocumento41 páginasClase 4 Unidad 3 - ReplicaciónLeilaAún no hay calificaciones
- Replicacion Del ADN 26.03.2023Documento12 páginasReplicacion Del ADN 26.03.2023GabrielaAún no hay calificaciones
- Proyectos 5tos 2023: Proyecto TODocumento22 páginasProyectos 5tos 2023: Proyecto TOSlandyAún no hay calificaciones
- Nucleo Celular - Replicacion 2021-IIDocumento28 páginasNucleo Celular - Replicacion 2021-IIKaren FelixAún no hay calificaciones
- BIOLOGIA-ADN (01-06-2021) TareaDocumento5 páginasBIOLOGIA-ADN (01-06-2021) TareaEmanuel HFAún no hay calificaciones
- Práctica 4Documento8 páginasPráctica 4Antonella Ortiz100% (1)
- Adn, ArnDocumento2 páginasAdn, ArnIsabella AladinoAún no hay calificaciones
- Evaluación TranscripciónDocumento2 páginasEvaluación TranscripciónDIANA CAROLINA POSADA LINARESAún no hay calificaciones
- Preguntas y EjerciciosDocumento3 páginasPreguntas y EjerciciosMilena CruzAún no hay calificaciones
- Ciclo intensivo Biología UNALMDocumento5 páginasCiclo intensivo Biología UNALMdayroAún no hay calificaciones
- 5 SeminarioDocumento2 páginas5 Seminariocrashito247Aún no hay calificaciones
- Dogma central de la biología molecularDocumento4 páginasDogma central de la biología molecularAndres FelipeAún no hay calificaciones
- Replicación: Síntesis de ADN: Biología Molecular y GenéticaDocumento34 páginasReplicación: Síntesis de ADN: Biología Molecular y GenéticaJosé CisternasAún no hay calificaciones
- Taller de Trascripcion y TraduccionDocumento7 páginasTaller de Trascripcion y TraduccionmcAún no hay calificaciones
- Actividad Complementaria Síntesis de ADNDocumento3 páginasActividad Complementaria Síntesis de ADNAndrea Paola Chaparro OrtizAún no hay calificaciones
- Cuestionario Unidad 4Documento15 páginasCuestionario Unidad 4muriel aravena makucAún no hay calificaciones
- Genética BacterianaDocumento55 páginasGenética BacterianaRoque JuncalAún no hay calificaciones
- Actividad - Genetica Molecular Noveno12Documento6 páginasActividad - Genetica Molecular Noveno12Alexandra Pedroza100% (1)
- C Sem10 BiologiaDocumento7 páginasC Sem10 BiologiaEG YasadiAún no hay calificaciones
- Segundo Bloque ThaliaDocumento6 páginasSegundo Bloque ThaliaMaria Fernanda SilvaAún no hay calificaciones
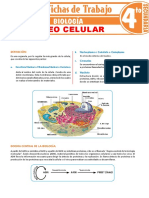
- Nucleo Celular para Cuarto Grado de SecundariaDocumento4 páginasNucleo Celular para Cuarto Grado de SecundariaGloria LorenzoAún no hay calificaciones
- Teoria Genetica MolecularDocumento153 páginasTeoria Genetica MolecularLucre CarnuccioAún no hay calificaciones
- Adn y ArnDocumento12 páginasAdn y Arnefrain juliaoAún no hay calificaciones
- Genética molecular: transcripción, replicación y expresión génicaDocumento9 páginasGenética molecular: transcripción, replicación y expresión génicaCardonaCristianAún no hay calificaciones
- Preicfes Nariño Con RespuestasDocumento9 páginasPreicfes Nariño Con RespuestasLuis Fernando Sanchez PucheAún no hay calificaciones
- Mendelclass Anatomia Celula 3 Nucleo y Ciclo Celular-17121020399Documento11 páginasMendelclass Anatomia Celula 3 Nucleo y Ciclo Celular-17121020399Claudio Galarzza FernándezAún no hay calificaciones
- GenéticaDocumento60 páginasGenéticaAguirre Gómez100% (1)
- Bmi TDocumento176 páginasBmi TJuan Arcos HuasasquicheAún no hay calificaciones
- Grupo 1 - Final Biología 15 - 02 - 2024Documento9 páginasGrupo 1 - Final Biología 15 - 02 - 2024themodmedbrAún no hay calificaciones
- PREICFES 9 Ciencias NaturalesDocumento3 páginasPREICFES 9 Ciencias NaturalesEstefania SalazarAún no hay calificaciones
- Genes, genoma y organización cromosómicaDocumento82 páginasGenes, genoma y organización cromosómicaDaniela TejadaAún no hay calificaciones
- Biología General - Semana Ix Expresión Génica - SemilleroDocumento5 páginasBiología General - Semana Ix Expresión Génica - SemilleroWilman IshpilcoAún no hay calificaciones
- Resumen Biologia CelularDocumento59 páginasResumen Biologia CelularCaru LitkeAún no hay calificaciones
- s3 Trascribiendo Genes en Plantas JDMMDocumento54 páginass3 Trascribiendo Genes en Plantas JDMMMax PlasenciaAún no hay calificaciones
- Clase 4. Replicacion Del ADNDocumento37 páginasClase 4. Replicacion Del ADNLuis Fabian Rosado PicazaAún no hay calificaciones
- Seminario BC 7-2020Documento68 páginasSeminario BC 7-2020Andrea Soledad PerezAún no hay calificaciones
- Cuestionario Sobre ADNDocumento1 páginaCuestionario Sobre ADNLibrofilo100% (2)
- Solemnes Biología MolecularDocumento12 páginasSolemnes Biología MolecularLeonidas LarrondoAún no hay calificaciones
- TALLER 04 Genetica Molecular 2017Documento2 páginasTALLER 04 Genetica Molecular 2017Taydith Teresa Sierra Tibamoza0% (1)
- Semana 10. Estructura Del ADN y ARN - GRADO 9Documento13 páginasSemana 10. Estructura Del ADN y ARN - GRADO 9Castro Abogados & AsesoresAún no hay calificaciones
- CROMATINADocumento3 páginasCROMATINASalomé EstévezAún no hay calificaciones
- Tema 2.1 Técnicas Citogenéticas y Moleculares.Documento6 páginasTema 2.1 Técnicas Citogenéticas y Moleculares.Cony CastañedaAún no hay calificaciones
- Acidos Nucleicos ADN y ARNDocumento3 páginasAcidos Nucleicos ADN y ARNLuis A. Romero0% (1)
- Evaluacion Sintesis de ProteinasDocumento1 páginaEvaluacion Sintesis de ProteinasRosiDanisAún no hay calificaciones
- SEMANA 10-Dogma CentralDocumento2 páginasSEMANA 10-Dogma CentralEduardo33% (3)
- Práctica 4 - Expresión Génica y Código GenéticoDocumento3 páginasPráctica 4 - Expresión Génica y Código GenéticoRuth ParedesAún no hay calificaciones
- Dogma Central 2018 EpigeneticaDocumento151 páginasDogma Central 2018 EpigeneticaOsmar Leonardo Gavilan SotoAún no hay calificaciones
- Expresión génica ANUAL UNIDocumento3 páginasExpresión génica ANUAL UNIDaniel TorresAún no hay calificaciones
- ACIDOS NUCLEICOS y TRANSMISION de La INFORMACION GENETICADocumento57 páginasACIDOS NUCLEICOS y TRANSMISION de La INFORMACION GENETICAJudith SheilaAún no hay calificaciones
- 4.1 Bases Físicas de La Herencia 1Documento25 páginas4.1 Bases Físicas de La Herencia 1Dario VelaAún no hay calificaciones
- Preguntas BiologiaDocumento6 páginasPreguntas BiologiaAndrea Pérez AriasAún no hay calificaciones
- Núcleo InterfásicoDocumento4 páginasNúcleo InterfásicoPatricia EncinaAún no hay calificaciones
- 6 - Mini Apunte Transcripción Y TraducciónDocumento12 páginas6 - Mini Apunte Transcripción Y TraducciónLeonardo BatistaAún no hay calificaciones
- Semana 07 - BiologíaDocumento4 páginasSemana 07 - BiologíaLuis Daniel Llontop TenorioAún no hay calificaciones
- Reconocimiento Unidad 2Documento6 páginasReconocimiento Unidad 2Manuel Ricardo Bayona PerezAún no hay calificaciones
- Semana 3.1-Dogma de La BiologiaaDocumento9 páginasSemana 3.1-Dogma de La Biologiaaaimipatogallina1234Aún no hay calificaciones
- MGB 1 - TeoríaDocumento32 páginasMGB 1 - TeoríaAgustin TuroneAún no hay calificaciones
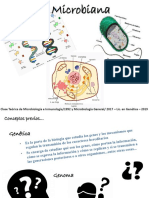
- Genética MicrobianaDocumento31 páginasGenética MicrobianaPaula DanielaAún no hay calificaciones
- REPLICACIONDocumento21 páginasREPLICACIONElias CortezAún no hay calificaciones
- Reaccion Quimica Diferente de MezclaDocumento3 páginasReaccion Quimica Diferente de MezclaAsasAún no hay calificaciones
- Lab Microbiología Bajo CostoDocumento2 páginasLab Microbiología Bajo CostoAsasAún no hay calificaciones
- 02 Stem Tecno WebDocumento24 páginas02 Stem Tecno WebAsasAún no hay calificaciones
- 5-La Vida Al Interior Del Genoma-EnviadoDocumento8 páginas5-La Vida Al Interior Del Genoma-EnviadoBRYAN CAMILO MUÑOZ AVILESAún no hay calificaciones
- CONTABILIDAD BÁSICA - Practico 06 Ejercicios Sobre Registración en Libro DiarioDocumento4 páginasCONTABILIDAD BÁSICA - Practico 06 Ejercicios Sobre Registración en Libro DiarioDiego Soto MoyaAún no hay calificaciones
- Plantilla - Presentación - Diagrama V de GowinDocumento4 páginasPlantilla - Presentación - Diagrama V de GowinyurleyAún no hay calificaciones
- Catálogo Final Remate Luis GuzmánDocumento68 páginasCatálogo Final Remate Luis GuzmánLopecito LpdtAún no hay calificaciones
- Devocional BíblicoDocumento16 páginasDevocional BíblicoMartin Antonio Santa Cruz QuevedoAún no hay calificaciones
- Tañido de Una FlautaDocumento2 páginasTañido de Una FlautaSergio Alberto Cortés RonquilloAún no hay calificaciones
- Test PsicológicosDocumento5 páginasTest PsicológicosFilomena Andrea GonzálezAún no hay calificaciones
- Del Sexo A La Super Conciencia - OshoDocumento27 páginasDel Sexo A La Super Conciencia - Oshomiryamrugo100% (1)
- Exp1 Eba IntermedioDocumento12 páginasExp1 Eba IntermedioSANDRA ArticaAún no hay calificaciones
- Bujia Arte Contemporaneo 1Documento42 páginasBujia Arte Contemporaneo 1Osvaldo Hernández MuroAún no hay calificaciones
- Iacc Control Segundo SemestreDocumento5 páginasIacc Control Segundo SemestreHANS KURTAún no hay calificaciones
- 18 Concreto Sin FinosDocumento5 páginas18 Concreto Sin FinosEloy A. Cabrera ZuñigaAún no hay calificaciones
- Bernaza, Cammotto y LongoDocumento20 páginasBernaza, Cammotto y LongoIsabel PérezAún no hay calificaciones
- 12 Piedras CementadasDocumento15 páginas12 Piedras CementadasCesar O. Retamar RomeroAún no hay calificaciones
- Practica 1Documento7 páginasPractica 1Edith R ChAún no hay calificaciones
- Manual de organización y funciones DEMEMDocumento66 páginasManual de organización y funciones DEMEMjomer stevenAún no hay calificaciones
- VRMMP Auditorías MortalidadDocumento4 páginasVRMMP Auditorías MortalidadJordany Placencia0% (1)
- Trabajo Final DinamicaDocumento8 páginasTrabajo Final DinamicaNorbil Correa gilAún no hay calificaciones
- CLASIFICACIÓN DE PARÁSITOS SEGÚN NIVEL DE BIOSEGURIDADDocumento1 páginaCLASIFICACIÓN DE PARÁSITOS SEGÚN NIVEL DE BIOSEGURIDADDiego Erazo GirónAún no hay calificaciones
- Poster PDFDocumento1 páginaPoster PDFEstaban Ossa100% (1)
- Puntos y Planos Cefalometricos (Word)Documento4 páginasPuntos y Planos Cefalometricos (Word)cnvelasquezAún no hay calificaciones
- Componentes Del Servicio Al ClienteDocumento1 páginaComponentes Del Servicio Al ClienteSonia Pascual ChavezAún no hay calificaciones
- La Obra de Juan Ferrando Badía y Su Significación en El Desarrollo Del Derecho Constitucional y de La Ciencia Política en EspañaDocumento38 páginasLa Obra de Juan Ferrando Badía y Su Significación en El Desarrollo Del Derecho Constitucional y de La Ciencia Política en EspañaRenzzo Paolo Retes PaucarAún no hay calificaciones
- Los Booms Petroleros ¿Qué Cambió en Los Últimos 40 Años Foro Economía EcuadorDocumento1 páginaLos Booms Petroleros ¿Qué Cambió en Los Últimos 40 Años Foro Economía EcuadorGame DylánAún no hay calificaciones
- Corrientes SociopolíticasDocumento9 páginasCorrientes SociopolíticasJair Exzacarias Hernández100% (2)
- Agenda Lunes 25 Al 29 de OctubreDocumento5 páginasAgenda Lunes 25 Al 29 de OctubreGisela PeñaAún no hay calificaciones
- Reg - Aux.consol 1T 2 A2 AritDocumento6 páginasReg - Aux.consol 1T 2 A2 AritElder Junior Roman LeonAún no hay calificaciones
- Desplazamiento MiscibleDocumento34 páginasDesplazamiento MisciblePedro Reynaldo Marin Dominguez100% (1)
- Portafolio. Nuevos Modelos de NegociosDocumento27 páginasPortafolio. Nuevos Modelos de NegociosVictor CavazosAún no hay calificaciones
- Anexo 1 - Colaborativa-238Documento10 páginasAnexo 1 - Colaborativa-238vanesa telloAún no hay calificaciones
- Responsabilidad Por La Función Pública BoliviaDocumento31 páginasResponsabilidad Por La Función Pública BoliviaSoriaVargas Edwin95% (21)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Genética general: Libro de textoDe EverandGenética general: Libro de textoCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Neuropsicología: Los fundamentos de la materiaDe EverandNeuropsicología: Los fundamentos de la materiaCalificación: 5 de 5 estrellas5/5 (1)
- Los cinco ritos tibetanos de la eterna juventud: El ojo de la revelaciónDe EverandLos cinco ritos tibetanos de la eterna juventud: El ojo de la revelaciónCalificación: 4.5 de 5 estrellas4.5/5 (12)
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- Hormonas: Mensajeros químicos y comunicación celularDe EverandHormonas: Mensajeros químicos y comunicación celularCalificación: 2.5 de 5 estrellas2.5/5 (2)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Minerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.De EverandMinerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.Calificación: 3 de 5 estrellas3/5 (2)
- Abrázame fuerte: Siete conversaciones para un amor duraderoDe EverandAbrázame fuerte: Siete conversaciones para un amor duraderoManu BerásteguiCalificación: 4.5 de 5 estrellas4.5/5 (13)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- El autismo: Reflexiones y pautas para comprenderlo y abordarloDe EverandEl autismo: Reflexiones y pautas para comprenderlo y abordarloCalificación: 4 de 5 estrellas4/5 (7)
- El perfil psicológico de Jesús: Aprendamos del Maestro a manejar efectivamente nuestras emocionesDe EverandEl perfil psicológico de Jesús: Aprendamos del Maestro a manejar efectivamente nuestras emocionesCalificación: 4 de 5 estrellas4/5 (4)
- Terapia de vidas pasadas: Un camino hacia la luz del alma. Técnica y prácticaDe EverandTerapia de vidas pasadas: Un camino hacia la luz del alma. Técnica y prácticaCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Bases biológicas del comportamiento animal y humanoDe EverandBases biológicas del comportamiento animal y humanoCalificación: 4 de 5 estrellas4/5 (4)
- La familia: Comprensión dinámica e intervenciones terapéuticasDe EverandLa familia: Comprensión dinámica e intervenciones terapéuticasCalificación: 5 de 5 estrellas5/5 (1)
- Nuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosDe EverandNuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosCalificación: 3.5 de 5 estrellas3.5/5 (6)