También podría gustarte
- Novena PracticaDocumento10 páginasNovena PracticaJonAún no hay calificaciones
- Cuestionario ClonaciónDocumento4 páginasCuestionario ClonaciónalejandraAún no hay calificaciones
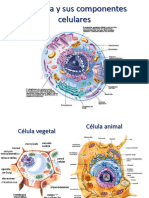
- Clase 1 La CelulaDocumento35 páginasClase 1 La Celularossmery briceño balmacedaAún no hay calificaciones
- Presentación Membrana PlasmáticaDocumento26 páginasPresentación Membrana PlasmáticaMiguel CastillaAún no hay calificaciones
- Limites Celulares.1Documento62 páginasLimites Celulares.1Libny Quispe SegoviaAún no hay calificaciones
- TEMA 2. Membrana BiológicaDocumento6 páginasTEMA 2. Membrana BiológicaLaura FernándezAún no hay calificaciones
- Unidad IIDocumento44 páginasUnidad IIruben.lv2330Aún no hay calificaciones
- Organelos CelularesDocumento77 páginasOrganelos CelularesSweetLadyAún no hay calificaciones
- 02 Membrana y Transporte CelularDocumento35 páginas02 Membrana y Transporte CelularAnonymous wakgFRKAún no hay calificaciones
- Membrana PlasmáticaDocumento28 páginasMembrana PlasmáticajannyyfcAún no hay calificaciones
- La Célula 1ºbachDocumento8 páginasLa Célula 1ºbachnhc8j2jbg4Aún no hay calificaciones
- Biologia Celular e Histologia PARTE1Documento38 páginasBiologia Celular e Histologia PARTE1Raquel DiezAún no hay calificaciones
- Membrana PlasmáticaDocumento18 páginasMembrana PlasmáticaInés RodríguezAún no hay calificaciones
- Tema 2 - Membrana Celular y CitoplasmaDocumento11 páginasTema 2 - Membrana Celular y CitoplasmaALETEPAún no hay calificaciones
- Esquema Resumen 7Documento15 páginasEsquema Resumen 7Jennifer Diaz PomaAún no hay calificaciones
- Membra NaDocumento33 páginasMembra NaGilmer Gustavo EzamAún no hay calificaciones
- 3 - Célula y Membrana CelularDocumento32 páginas3 - Célula y Membrana Celularjose.torres1Aún no hay calificaciones
- 02 La CelulaDocumento51 páginas02 La CelulaAndrea MaríaAún no hay calificaciones
- Membrana y TransprteDocumento42 páginasMembrana y Transprtemaria alejandra martinez sanabriaAún no hay calificaciones
- Membrana Celular Estructura Funcion yDocumento82 páginasMembrana Celular Estructura Funcion yRafaelCamargoArguellesAún no hay calificaciones
- Nivel Tisular de OrganizacionDocumento45 páginasNivel Tisular de OrganizacionVanessa VargasAún no hay calificaciones
- Membrana Celular y TransporteDocumento9 páginasMembrana Celular y TransporteAlejandraAún no hay calificaciones
- 1.2.3 Célula VegetalDocumento32 páginas1.2.3 Célula VegetalSonia Herrera MonroyAún no hay calificaciones
- Apuntes FisiologíaDocumento272 páginasApuntes FisiologíaIsabel Vivas JaramilloAún no hay calificaciones
- Cel EucariotaDocumento49 páginasCel Eucariotanive cuestaAún no hay calificaciones
- Membrana Citroplasmática 28-10-2020Documento23 páginasMembrana Citroplasmática 28-10-2020Eddy Gabriel OlmedoAún no hay calificaciones
- 9-1-La Membrana Plasmatica y Otros OrganulosDocumento42 páginas9-1-La Membrana Plasmatica y Otros OrganulosDaniel IbayAún no hay calificaciones
- Membranas Biológicas - OML 2023ADocumento30 páginasMembranas Biológicas - OML 2023ADhara RicciAún no hay calificaciones
- Membrana Celular 15-09aDocumento80 páginasMembrana Celular 15-09aricardo medina sulcaAún no hay calificaciones
- Nivel Cel CompletoDocumento55 páginasNivel Cel Completonayelicahue8Aún no hay calificaciones
- 1-Introducción A La Fisiología.Documento71 páginas1-Introducción A La Fisiología.jgarciasAún no hay calificaciones
- Copia de Guía de Laboratorio 3Documento36 páginasCopia de Guía de Laboratorio 3sara barriosAún no hay calificaciones
- Celula 2022Documento60 páginasCelula 2022Meli ParedesAún no hay calificaciones
- Biología Clase 13-14Documento29 páginasBiología Clase 13-14ASMAún no hay calificaciones
- TEMA 2 - La Membrana PlasmáticaDocumento8 páginasTEMA 2 - La Membrana PlasmáticaIlAguacherry IlAún no hay calificaciones
- Sesión 04 - Biología CelularDocumento40 páginasSesión 04 - Biología CelularSILVINA JUDITH CABRERA MATTOSAún no hay calificaciones
- RESUMENES DE TEORIA - BIOLOGÍA GENERAL Y CELULAR 1rDocumento26 páginasRESUMENES DE TEORIA - BIOLOGÍA GENERAL Y CELULAR 1rroxcab003Aún no hay calificaciones
- Resumen de HistologiaDocumento45 páginasResumen de HistologiaRomina CasanovaAún no hay calificaciones
- Clase 4 BH Unidad 2Documento8 páginasClase 4 BH Unidad 2Josefa 001Aún no hay calificaciones
- Membranas CelularesDocumento4 páginasMembranas CelularesDaniela MartinezAún no hay calificaciones
- Tema 4. La Estructura de La CélulaDocumento6 páginasTema 4. La Estructura de La CélulaPablo Martos AguayoAún no hay calificaciones
- Membrana CelularDocumento16 páginasMembrana CelularJulio Luna100% (1)
- Organelos y CelulasDocumento30 páginasOrganelos y CelulasRomilioAntonioRetamalesGermanottaEncinaAún no hay calificaciones
- Guía Fisiología - Celular y RespiratorioDocumento51 páginasGuía Fisiología - Celular y RespiratorioMarines BarretoAún no hay calificaciones
- MEMBRANASDocumento70 páginasMEMBRANASDiana Vite RomeroAún no hay calificaciones
- Biomoléculas Membranas - Ing SaludDocumento3 páginasBiomoléculas Membranas - Ing SaludLucia Jimenez VegaAún no hay calificaciones
- Biologia CelularDocumento13 páginasBiologia CelulargabrielaaAún no hay calificaciones
- LA CELULA CUARTA A Y B y TrabajoDocumento25 páginasLA CELULA CUARTA A Y B y Trabajobasilia quispe incahuanacoAún no hay calificaciones
- Capítulo 3 - CitoplasmaDocumento38 páginasCapítulo 3 - CitoplasmaKaren Velázquez BravoAún no hay calificaciones
- Membrana CelularDocumento58 páginasMembrana CelularAldo AcostaAún no hay calificaciones
- CitoplasmaDocumento30 páginasCitoplasmaKeny Mamani DuriAún no hay calificaciones
- Tema 7. Las Envolturas CelularesDocumento12 páginasTema 7. Las Envolturas CelularesMarco MatellánAún no hay calificaciones
- Membrana y Transporte PDFDocumento48 páginasMembrana y Transporte PDFMarielos VelásquezAún no hay calificaciones
- Membrana CelularDocumento22 páginasMembrana CelularManuel Cespedes100% (1)
- TEMA2 BiocelDocumento23 páginasTEMA2 BiocelMóni GonzálezAún no hay calificaciones
- U.D.1.1.1 O. Microscopica (La Célula)Documento15 páginasU.D.1.1.1 O. Microscopica (La Célula)Adriel Rodriguez AgudoAún no hay calificaciones
- 1 Las Células Mitosis MeiosisDocumento104 páginas1 Las Células Mitosis MeiosismateohernandezAún no hay calificaciones
- Histología - MODULO BDocumento6 páginasHistología - MODULO BXimin XaoAún no hay calificaciones
- Fisiología CelularDocumento2 páginasFisiología CelularEduardo Jahir AnguloAún no hay calificaciones
- Celula VegetalDocumento11 páginasCelula VegetalBarbara ReyesAún no hay calificaciones
- Semana 5 Teoria Fundamentos Biologicos-IDocumento60 páginasSemana 5 Teoria Fundamentos Biologicos-IAstrid GallegosAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- 687 2564 1 PBDocumento12 páginas687 2564 1 PBLinbert Simon CallataAún no hay calificaciones
- GLOSARIODocumento9 páginasGLOSARIOJork GaddeaAún no hay calificaciones
- F-HC-47 Consentimiento Informado para Prueba de Secuenciación de Exoma Clínico - GenomaCESDocumento4 páginasF-HC-47 Consentimiento Informado para Prueba de Secuenciación de Exoma Clínico - GenomaCESalejandro restrepoAún no hay calificaciones
- Introducc GenmolDocumento45 páginasIntroducc GenmolJosue EscarcegaAún no hay calificaciones
- Genoma HumanoDocumento27 páginasGenoma HumanoLaly QuintanaAún no hay calificaciones
- Síndrome de Pierre-RobinDocumento15 páginasSíndrome de Pierre-RobinPaulina Santana100% (1)
- Virus - Un Error de Interpretación (Parte II) - MaterialdenmgDocumento14 páginasVirus - Un Error de Interpretación (Parte II) - MaterialdenmgLiza MarieAún no hay calificaciones
- Manual Pract GIIDocumento148 páginasManual Pract GIIromerilloAún no hay calificaciones
- Pac BioDocumento13 páginasPac BioJanethGutierrezDiazAún no hay calificaciones
- Parcial GeneticaDocumento54 páginasParcial GeneticaRaquel Pérez FernándezAún no hay calificaciones
- Génetica Micro. UNIDAD 11 O - 2020Documento44 páginasGénetica Micro. UNIDAD 11 O - 2020Ariadna Mariel MoralesAún no hay calificaciones
- Ingeniería GenéticaDocumento34 páginasIngeniería GenéticaJairo CardenasAún no hay calificaciones
- Electroforesis en Gel de Agarosa - Fernandez Salazar Iris AzucenaDocumento18 páginasElectroforesis en Gel de Agarosa - Fernandez Salazar Iris AzucenaIris Azucena Fernández SalazarAún no hay calificaciones
- Guía de Prácticas Pinares 2022Documento29 páginasGuía de Prácticas Pinares 2022Ruben FuentesAún no hay calificaciones
- Evaluación Correspondiente Al: Telesecundaria 1° GradoDocumento19 páginasEvaluación Correspondiente Al: Telesecundaria 1° GradoLorena Plata RomanAún no hay calificaciones
- Adn e Ingenieria GeneticaDocumento37 páginasAdn e Ingenieria GeneticaAdolfo GonzalezAún no hay calificaciones
- 2.03.02. Bronquitis Infecciosa Aviar PDFDocumento15 páginas2.03.02. Bronquitis Infecciosa Aviar PDFSaul Utani CayllahuaAún no hay calificaciones
- Apuntes BiotecnologiaDocumento38 páginasApuntes BiotecnologiaStevenAún no hay calificaciones
- 2.1.04 Viremia Primaveral de La CarpaDocumento14 páginas2.1.04 Viremia Primaveral de La CarpaguvalAún no hay calificaciones
- Lipasas PapeleriaDocumento5 páginasLipasas PapeleriaAntonio Marques GutierrezAún no hay calificaciones
- Secuenciación de Contenidos de ArtesDocumento30 páginasSecuenciación de Contenidos de ArtesOsmar Espinosa CigarroaAún no hay calificaciones
- Entrega Final Scheduling e InventariosDocumento19 páginasEntrega Final Scheduling e InventariosJohan Diaz75% (4)
- Primer DegeneradoDocumento9 páginasPrimer DegeneradoSandra MolinaAún no hay calificaciones
- Eco-Epidemiología de Garrapatas y Rickettsias en El Trópico ColombianoDocumento24 páginasEco-Epidemiología de Garrapatas y Rickettsias en El Trópico ColombianoEfrain BenavidesAún no hay calificaciones
- Técnicas de Detección de PolimorfismosDocumento24 páginasTécnicas de Detección de PolimorfismosXael GarciaAún no hay calificaciones
- Tema 7Documento24 páginasTema 7Laura Saez TinaqueroAún no hay calificaciones
- Guía PCR - 2022-1Documento4 páginasGuía PCR - 2022-1Mari PeñaAún no hay calificaciones
- Proyecto LitografiaDocumento12 páginasProyecto LitografiaCarlos Pinzon100% (1)