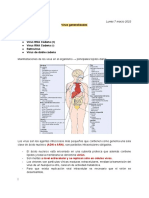
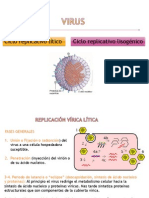
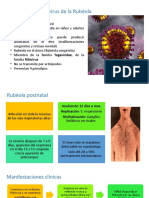
También podría gustarte
- VIROLOGIADocumento6 páginasVIROLOGIAtatiana naranjoAún no hay calificaciones
- Genetica-Virus-Pato e InmunidadDocumento63 páginasGenetica-Virus-Pato e InmunidadOriana MuruaAún no hay calificaciones
- MicrobiologíaDocumento139 páginasMicrobiologíaale_mc98Aún no hay calificaciones
- Clasificación y estructura de los virusDocumento4 páginasClasificación y estructura de los viruskimberlynAún no hay calificaciones
- Virus, Hongos y Parasitos MicroDocumento67 páginasVirus, Hongos y Parasitos MicroDebob16Aún no hay calificaciones
- VIROLOGÍA Clase I - III 18 PDFDocumento108 páginasVIROLOGÍA Clase I - III 18 PDFKarol Verenisse Gamarra BustamanteAún no hay calificaciones
- Exposicion VirusDocumento43 páginasExposicion VirusEduardo GarciaAún no hay calificaciones
- Virus: estructura, características y clasificaciónDocumento32 páginasVirus: estructura, características y clasificaciónjhony100% (1)
- Iv ComplementoDocumento4 páginasIv ComplementoLUIS DAVID PEREZ MERCADOAún no hay calificaciones
- Biologia ViralDocumento6 páginasBiologia ViralSerenaAún no hay calificaciones
- Guia 8 Virus-Sida Biologia Semana 8Documento17 páginasGuia 8 Virus-Sida Biologia Semana 8Jamile Barrientos SánchezAún no hay calificaciones
- Virus Resumen SelectividadDocumento3 páginasVirus Resumen SelectividadOsvaldo Elizalde ArmasAún no hay calificaciones
- Clasificación de BaltimoreDocumento3 páginasClasificación de BaltimoreAngela OrietaAún no hay calificaciones
- Ensayo de Adn y ArnDocumento7 páginasEnsayo de Adn y ArnMassiel Argentina Zeledon CaleroAún no hay calificaciones
- TEO 1073 Sesión 5 Generalidades de VirusDocumento21 páginasTEO 1073 Sesión 5 Generalidades de VirusarteagadannnAún no hay calificaciones
- VIROLOGIADocumento8 páginasVIROLOGIAKAREN ORELLANAAún no hay calificaciones
- Semana 2 VIRUSDocumento4 páginasSemana 2 VIRUSMaria GutierrezAún no hay calificaciones
- Resumen de VirusDocumento7 páginasResumen de VirusMaty EspinosaAún no hay calificaciones
- Características y clasificación de virusDocumento17 páginasCaracterísticas y clasificación de virusOriana MuruaAún no hay calificaciones
- Resuemen de VirologiaDocumento79 páginasResuemen de VirologiaCésar AlcaláAún no hay calificaciones
- Guía Verano SM N°1 (CB)Documento8 páginasGuía Verano SM N°1 (CB)Evelin Cueva ValverdeAún no hay calificaciones
- Triptico Los VirusDocumento25 páginasTriptico Los VirusArianny AcasmeAún no hay calificaciones
- Generalidades de Virus 2022-1Documento22 páginasGeneralidades de Virus 2022-1Juanes Villa100% (1)
- TEMA 18Documento2 páginasTEMA 18Rosa Alonso-VillalobosAún no hay calificaciones
- Virus AdnDocumento13 páginasVirus AdnDIANA GABRIELA TORRES OLIVERAAún no hay calificaciones
- Clase 5 - VirusDocumento22 páginasClase 5 - VirusKatAún no hay calificaciones
- Clasificación de BaltimoreDocumento4 páginasClasificación de BaltimoreAnny Arizmendi100% (1)
- VIRUS Parte 1Documento136 páginasVIRUS Parte 1Grecia González OcampoAún no hay calificaciones
- 5 VirusDocumento45 páginas5 VirusMoskito SchneiderAún no hay calificaciones
- Resumen VirusDocumento5 páginasResumen VirusNattsAún no hay calificaciones
- Virus PDFDocumento76 páginasVirus PDFRocioAún no hay calificaciones
- Clases de virus según su material genético y replicaciónDocumento4 páginasClases de virus según su material genético y replicaciónAnthony SuarezAún no hay calificaciones
- Propiedades químicas de los virus: ácidos nucleicos y clasificación de BaltimoreDocumento30 páginasPropiedades químicas de los virus: ácidos nucleicos y clasificación de BaltimoreCarlos Gerardo Castillo SosaAún no hay calificaciones
- VirusDocumento6 páginasVirusgb5sjkyj65Aún no hay calificaciones
- Tema 9Documento16 páginasTema 9elizabeth sarahy maggiorani cegarraAún no hay calificaciones
- Tema 1 Introduccion A La Virologia CompletoDocumento8 páginasTema 1 Introduccion A La Virologia CompletoDANIEL MANCILLA GABRIELAún no hay calificaciones
- 2 - Riesgos Biologos Virus Protozoss AcDocumento42 páginas2 - Riesgos Biologos Virus Protozoss AcLyzbeth LyzAún no hay calificaciones
- VirusDocumento13 páginasVirusLuz Anchiraico ramiresAún no hay calificaciones
- Historia y estructura de los virus enDocumento31 páginasHistoria y estructura de los virus enItandehui Jerónimo VeranoAún no hay calificaciones
- Clasificacion de Los Virus Según BaltimoreDocumento2 páginasClasificacion de Los Virus Según BaltimorearelyscaroliinaAún no hay calificaciones
- VirusDocumento41 páginasVirusJhon Alexis M ArgoteAún no hay calificaciones
- Virus y Hepatitis - Docx-2Documento18 páginasVirus y Hepatitis - Docx-2Daniela Ivanova Alvarado InfanteAún no hay calificaciones
- Apuntes Virología Completos Todas Las ClasesDocumento369 páginasApuntes Virología Completos Todas Las Clasespogah89776Aún no hay calificaciones
- Microbiología Unidad 5 FINALDocumento32 páginasMicrobiología Unidad 5 FINALJoaquínA.GallardoAún no hay calificaciones
- Replicación de Los VirusDocumento22 páginasReplicación de Los VirusLuis Fernando Avila TobiasAún no hay calificaciones
- Manual de Agentes Vivos de EnfermedadDocumento186 páginasManual de Agentes Vivos de EnfermedadSeba VielmaAún no hay calificaciones
- Estructura de Los VirusDocumento69 páginasEstructura de Los Virusjoser4foreverAún no hay calificaciones
- Clase de Virus FinalDocumento42 páginasClase de Virus FinalMelisa NuñezAún no hay calificaciones
- Virus ARN BicatenarioDocumento4 páginasVirus ARN BicatenarioRodrigo ReneAún no hay calificaciones
- Objetivos GeneralesDocumento5 páginasObjetivos GeneralesmartovpAún no hay calificaciones
- Manual AVE PDFDocumento178 páginasManual AVE PDFFrancisco martinezAún no hay calificaciones
- VIRUSDocumento3 páginasVIRUSPaul AguaisaAún no hay calificaciones
- VirusDocumento20 páginasVirusIngrid HernándezAún no hay calificaciones
- Genoma ViricoDocumento7 páginasGenoma ViricoSaul Adrian Mamani SanchezAún no hay calificaciones
- Guia Teoria VirusDocumento38 páginasGuia Teoria VirusVictor CetchaAún no hay calificaciones
- Los Virus (Apuntes)Documento8 páginasLos Virus (Apuntes)Ezequiel GuardiaAún no hay calificaciones
- Micro Biologia Togaviridae y ArteviridaeDocumento4 páginasMicro Biologia Togaviridae y ArteviridaeRusselAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- PROTOCOLO DE MUFACE PARA REALIZACIÓN DE PCRs AL CNPDocumento2 páginasPROTOCOLO DE MUFACE PARA REALIZACIÓN DE PCRs AL CNPufpalmeriaAún no hay calificaciones
- Autodiagnostico Medidas Covid 19 para EmpresasDocumento3 páginasAutodiagnostico Medidas Covid 19 para EmpresasVivi Hernandez RubilarAún no hay calificaciones
- COVID 19, LA PANDEMIA POR CORONAVIRUS - Enfrentando A Un Enemigo Invisible (Spanish Edition)Documento220 páginasCOVID 19, LA PANDEMIA POR CORONAVIRUS - Enfrentando A Un Enemigo Invisible (Spanish Edition)artAún no hay calificaciones
- Monografia ImunologicoDocumento19 páginasMonografia ImunologicoLeydi Kiara Silva LopezAún no hay calificaciones
- TRS MC2Documento2 páginasTRS MC2BERNIE LEANDRO PUJADAS CONTRERASAún no hay calificaciones
- Vacuna Contra La COVID V1Documento8 páginasVacuna Contra La COVID V1William DextreAún no hay calificaciones
- EVALUACIONESDocumento12 páginasEVALUACIONESDenisAún no hay calificaciones
- Aislamiento social por Covid-19Documento3 páginasAislamiento social por Covid-19Arphi Juan50% (2)
- Cat Wondfo+doraDocumento36 páginasCat Wondfo+doraElizabeth RiveraAún no hay calificaciones
- Inmunodeficiencia de Células T Con Epidermodisplasia VerruciformeDocumento5 páginasInmunodeficiencia de Células T Con Epidermodisplasia VerruciformeEliana Gómez ReyesAún no hay calificaciones
- Mapa Conceptual de BacterDocumento1 páginaMapa Conceptual de BacterCervantes Sánchez Alma JazmínAún no hay calificaciones
- Ficha de La Ada1 - 2do - Sistema RespiratorioDocumento4 páginasFicha de La Ada1 - 2do - Sistema RespiratorioNicanor Velásquez AcuñaAún no hay calificaciones
- Niveles de Riesgo - COVID19 - DITENDocumento15 páginasNiveles de Riesgo - COVID19 - DITENhenryAún no hay calificaciones
- Protocolo para Establecimientos Gastronomicos 06jun20 3Documento18 páginasProtocolo para Establecimientos Gastronomicos 06jun20 3Cadena 3 Argentina100% (1)
- Unidad 1 SicologiaDocumento7 páginasUnidad 1 SicologiaAna GonzalezAún no hay calificaciones
- Ecología microbiota bucalDocumento14 páginasEcología microbiota bucalOrme TexAún no hay calificaciones
- Discusion Informe 8Documento6 páginasDiscusion Informe 8Daniel Romero GarciaAún no hay calificaciones
- RubéolaDocumento10 páginasRubéolaHsjsisAún no hay calificaciones
- Qué Es El VIHDocumento3 páginasQué Es El VIHgreinyerAún no hay calificaciones
- Examen Final-Comunicación y ArgumentaciónDocumento4 páginasExamen Final-Comunicación y Argumentacióndomrickypr100% (2)
- SARS3Documento22 páginasSARS3Natalia Martínez HernándezAún no hay calificaciones
- Herpes Zoster: Introdução e Formulacao do ProblemaDocumento3 páginasHerpes Zoster: Introdução e Formulacao do ProblemaGracianoAún no hay calificaciones
- Examenes Pasados Finales Curso BasicoDocumento98 páginasExamenes Pasados Finales Curso BasicoMaria SanchezAún no hay calificaciones
- Protocolo de Medidas Preventivas Frente Al Coronavirus-SERVICIOS SUBMARINOS ROMAN ORDOÑEZ E.I.R.L.Documento3 páginasProtocolo de Medidas Preventivas Frente Al Coronavirus-SERVICIOS SUBMARINOS ROMAN ORDOÑEZ E.I.R.L.Antony PumaAún no hay calificaciones
- Vacunas en Adultos MayoresDocumento7 páginasVacunas en Adultos MayoresAlejandra ZamudioAún no hay calificaciones
- La Sangre de Mi HijoDocumento15 páginasLa Sangre de Mi Hijobeto319Aún no hay calificaciones
- Algoritmo DX y Manejo Encefalitis.Documento3 páginasAlgoritmo DX y Manejo Encefalitis.diego riveraAún no hay calificaciones
- COVID-19 PerúDocumento3 páginasCOVID-19 PerúJuanPa ChanduvíAún no hay calificaciones
- Exantema SúbitoDocumento3 páginasExantema SúbitoKaReN YañezAún no hay calificaciones
- CancionesDocumento20 páginasCancionesRostuar OrloAún no hay calificaciones