También podría gustarte
- Histotecnologia GuiaDocumento9 páginasHistotecnologia GuiaMaría Fernanda YajuresAún no hay calificaciones
- La Educación Socialista en MéxicoDocumento17 páginasLa Educación Socialista en MéxicoEmulsión De Scott100% (40)
- Los Viajes de Una Gota de AguaDocumento2 páginasLos Viajes de Una Gota de AguaAdriana100% (1)
- Influencia de Un Campo Magnético Débil en El Comportamiento de Diadema Aff. Antillarum (Echinodermata: Echinoidea)Documento6 páginasInfluencia de Un Campo Magnético Débil en El Comportamiento de Diadema Aff. Antillarum (Echinodermata: Echinoidea)Paulina MoralesAún no hay calificaciones
- Guía 4 (Paleontología) - de Ávila, Jiménez, Mendoza y VesgaDocumento19 páginasGuía 4 (Paleontología) - de Ávila, Jiménez, Mendoza y VesgaJiménez 1408Aún no hay calificaciones
- Pistas FósilesDocumento10 páginasPistas FósilesElena ErasmusAún no hay calificaciones
- Estudio Comparativo de Biolocalización Entre Delfines y MurciélagosDocumento15 páginasEstudio Comparativo de Biolocalización Entre Delfines y MurciélagosJavier J SalazarAún no hay calificaciones
- Identificacion y Observacion de Protozoos Cultivados en Agua Estancada en Cartagena de IndiasDocumento3 páginasIdentificacion y Observacion de Protozoos Cultivados en Agua Estancada en Cartagena de IndiasMario Ali Lopez OrozcoAún no hay calificaciones
- Tipos de AdaptacionDocumento14 páginasTipos de AdaptacionArelys Mesa SanchezAún no hay calificaciones
- Identificación y Observación de Protozoos Cultivados en Agua Estancada en Cartagena de IndiasDocumento3 páginasIdentificación y Observación de Protozoos Cultivados en Agua Estancada en Cartagena de IndiasBRAYAN JOSE ORTEGA RICARDO ESTUDIANTEAún no hay calificaciones
- BIOFISICADocumento15 páginasBIOFISICAEfrain Ricardo Machado CampilloAún no hay calificaciones
- La Teoría de Biogeografia de Islas, Metapoblaciones, Modelacion de Nichos.Documento3 páginasLa Teoría de Biogeografia de Islas, Metapoblaciones, Modelacion de Nichos.Mara Paola Durango100% (1)
- Secuencia Didáctica NaturalesDocumento11 páginasSecuencia Didáctica NaturalesgisecappeAún no hay calificaciones
- Informe AnelidosDocumento5 páginasInforme Anelidosliz dayanna0% (1)
- 1.1. Evolución y Clasificación de Los Seres Vivos 22-23Documento62 páginas1.1. Evolución y Clasificación de Los Seres Vivos 22-23NeoCharlieAún no hay calificaciones
- Copie de DAMA DE ELCHEDocumento5 páginasCopie de DAMA DE ELCHEAlba Piedrabuena MartinezAún no hay calificaciones
- Tema 1. IntroducciónDocumento10 páginasTema 1. IntroducciónLucero TorradoAún no hay calificaciones
- Entomologia AplicadaDocumento326 páginasEntomologia AplicadaKarina De Avila AguirreAún no hay calificaciones
- ANÉLIDOSDocumento2 páginasANÉLIDOSpercy100% (1)
- Parcial 3 Parte 3Documento8 páginasParcial 3 Parte 3mariaamachucamAún no hay calificaciones
- Las Adaptaciones de Los Seres Vivos Al AmbienteDocumento6 páginasLas Adaptaciones de Los Seres Vivos Al Ambientenaychu10Aún no hay calificaciones
- Hipótesis Sobre Las Adaptaciones de Los Peces Payaso Amphiprion Percula y A. Ocellaris en Su Relación Simbiótica Con Las AnémonasDocumento18 páginasHipótesis Sobre Las Adaptaciones de Los Peces Payaso Amphiprion Percula y A. Ocellaris en Su Relación Simbiótica Con Las AnémonasBernardo Alfonso Fajardo Labra100% (1)
- Tarea BiologíaDocumento5 páginasTarea BiologíaMaria Victoria Altuv RanglAún no hay calificaciones
- Patrones de Evidencia Darwin y WallaceDocumento7 páginasPatrones de Evidencia Darwin y WallacedeathgamerAún no hay calificaciones
- Adaptación de Los Seres VivosDocumento31 páginasAdaptación de Los Seres Vivosapi-1962180783% (6)
- Biologia Segundo Medio Clase 2 Evidencias Evolutivas Parte IIDocumento10 páginasBiologia Segundo Medio Clase 2 Evidencias Evolutivas Parte IIDominique bucareyAún no hay calificaciones
- Nematodos 2 Rodríguez Rojas José AntonioDocumento6 páginasNematodos 2 Rodríguez Rojas José AntonioJose Antonio Rodriguez RojasAún no hay calificaciones
- Desarrollo Del Conocimientyo en El Estudio Del Comportamiento AnimalDocumento1 páginaDesarrollo Del Conocimientyo en El Estudio Del Comportamiento AnimalJEFFERSON DANIEL MONTENEGRO GONZALEZ (:-)Aún no hay calificaciones
- PoriferaDocumento19 páginasPoriferaIris MijaresAún no hay calificaciones
- Radiación AdaptativaDocumento5 páginasRadiación AdaptativaYimi BarraAún no hay calificaciones
- Trabajo Práctico Fotosintesis Parte 1Documento2 páginasTrabajo Práctico Fotosintesis Parte 1Roxana ChoqueAún no hay calificaciones
- El Sistema Nervioso, Clave de La EvoluciónDocumento4 páginasEl Sistema Nervioso, Clave de La EvoluciónBRENDA ALISON AGUILAR PABONAún no hay calificaciones
- PoriferosDocumento3 páginasPoriferosYessenia Ocsa SerranoAún no hay calificaciones
- ICFES Biologia 9 Mayo 1docxDocumento3 páginasICFES Biologia 9 Mayo 1docxCarolina andrea quintero TrianaAún no hay calificaciones
- Invertebrados de HumedalesDocumento2 páginasInvertebrados de HumedalesKalendra MiniaturasAún no hay calificaciones
- Nicho Conceptos y Radiacion AdapDocumento21 páginasNicho Conceptos y Radiacion AdapEliana Lopez FuentesAún no hay calificaciones
- Guia Teorias Evolutivas y Pruebas Ev Primero 2020Documento4 páginasGuia Teorias Evolutivas y Pruebas Ev Primero 2020jodoAún no hay calificaciones
- 5 PoriferaDocumento19 páginas5 PoriferaMIGUEL ALEX PEDRAZA ARANDOAún no hay calificaciones
- Actividad Sobre Video de El Origen de Las Especies Construyendo Una TeoríaDocumento3 páginasActividad Sobre Video de El Origen de Las Especies Construyendo Una TeoríaEspinoza Basurto Roberto CarlosAún no hay calificaciones
- Ficha 15. Explicaciones de Darwin Sobre Los Seres VivosDocumento8 páginasFicha 15. Explicaciones de Darwin Sobre Los Seres Vivosjose eduardo rodriguez zarateAún no hay calificaciones
- Práctico N°6 - Platyhelminthes y NemertiniDocumento11 páginasPráctico N°6 - Platyhelminthes y NemertiniCristian Santander RosalesAún no hay calificaciones
- ECOLOCALIZACIONDocumento3 páginasECOLOCALIZACIONoficinetAún no hay calificaciones
- Adaptaciones de Los Seres VivosDocumento7 páginasAdaptaciones de Los Seres Vivosfacavero96% (133)
- A L L (H) : TesisDocumento8 páginasA L L (H) : TesisGaston LezcanoAún no hay calificaciones
- 13 OrigEvoDomMamifChileDocumento18 páginas13 OrigEvoDomMamifChileclaudiaAún no hay calificaciones
- Teoria Evolutivas (Resumen) 9 de AgostoDocumento13 páginasTeoria Evolutivas (Resumen) 9 de Agostonailuj gonzalezAún no hay calificaciones
- Cuadro ComparativoDocumento5 páginasCuadro ComparativoDIANA MARCELA GONZALEZ HOYOSAún no hay calificaciones
- Biología de CordadosDocumento163 páginasBiología de CordadosNazareth ChávezAún no hay calificaciones
- Genaralidades PoríferosDocumento19 páginasGenaralidades PoríferosMiguel CalderonAún no hay calificaciones
- DragonfyDocumento7 páginasDragonfyValeria ArvalloAún no hay calificaciones
- Clase 8 Recolección de Muestras Biológicas para Identificación e Especies Nativas Traslado y Liberación.Documento40 páginasClase 8 Recolección de Muestras Biológicas para Identificación e Especies Nativas Traslado y Liberación.Maria Jose AsenjoAún no hay calificaciones
- Biología 7 Solucion Taller 1 PuntoDocumento6 páginasBiología 7 Solucion Taller 1 PuntoyeniferAún no hay calificaciones
- Infografia de Psicologia SocialDocumento1 páginaInfografia de Psicologia SocialAraceli Magaña PalmaAún no hay calificaciones
- Informe Anatomia de PecesDocumento2 páginasInforme Anatomia de PecesHangarAún no hay calificaciones
- Sistema de IntegraciónDocumento14 páginasSistema de IntegraciónDaniel WolfangAún no hay calificaciones
- Articulo Como Nemo Encontro Su HogarDocumento50 páginasArticulo Como Nemo Encontro Su HogarCami Becerra RodriguezAún no hay calificaciones
- Seminario Traducido 2.0Documento29 páginasSeminario Traducido 2.0Aldair Javier Ormeño AlcarrazAún no hay calificaciones
- Laboratorio#2 PaleontologíaDocumento11 páginasLaboratorio#2 PaleontologíaJuan Sebastian Càrdenas GuzmànAún no hay calificaciones
- Nematodos 1 Rodríguez Rojas José AntonioDocumento6 páginasNematodos 1 Rodríguez Rojas José AntonioJose Antonio Rodriguez RojasAún no hay calificaciones
- Algas de Yura ArequipaDocumento6 páginasAlgas de Yura Arequipaerick torresAún no hay calificaciones
- GRP BiodiversidadyEvoluciondeCnidarios Pena, ALDocumento2 páginasGRP BiodiversidadyEvoluciondeCnidarios Pena, ALMaría Fernanda YajuresAún no hay calificaciones
- AnemonasDocumento15 páginasAnemonasMaría Fernanda YajuresAún no hay calificaciones
- 397385991-Ordenanza-Simbolos-del-Municpio-san-Joaquin-Carabobo (Autoguardado)Documento3 páginas397385991-Ordenanza-Simbolos-del-Municpio-san-Joaquin-Carabobo (Autoguardado)María Fernanda YajuresAún no hay calificaciones
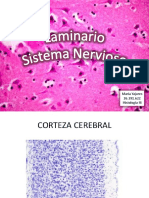
- LAMINARIO Sistema Nervioso HISTOLOGIADocumento36 páginasLAMINARIO Sistema Nervioso HISTOLOGIAMaría Fernanda YajuresAún no hay calificaciones
- Prohibido EnamorarseDocumento345 páginasProhibido EnamorarseMarylin YureyneAún no hay calificaciones
- Ocupación Territorial ChileDocumento3 páginasOcupación Territorial Chileingrid_vargas_hAún no hay calificaciones
- Actividades Bloques PoligonalesDocumento4 páginasActividades Bloques PoligonalesLeydi Betancourth Mesa100% (1)
- Resumen de Reconstrucción DisciplinarDocumento1 páginaResumen de Reconstrucción DisciplinarSandra RiquelmeAún no hay calificaciones
- La Oración Cristiana PDFDocumento5 páginasLa Oración Cristiana PDFAnthony SantillánAún no hay calificaciones
- Practica 1-Expo-Analisis de SacarosaDocumento9 páginasPractica 1-Expo-Analisis de SacarosaJuan Pablo Silvera MamaniAún no hay calificaciones
- La Sorpresa. MarcelliDocumento21 páginasLa Sorpresa. MarcelliGuillermina FernándezAún no hay calificaciones
- Incendia Mi AlmaDocumento1 páginaIncendia Mi AlmaLili RiveraAún no hay calificaciones
- Higiene y Seguridad IndustrialDocumento4 páginasHigiene y Seguridad IndustrialAnonymous 8H0pr5Aún no hay calificaciones
- Mendigo Ladron Guerrero y ReyDocumento11 páginasMendigo Ladron Guerrero y ReyRaulAlvarezAún no hay calificaciones
- El Abrazo de OsoDocumento1 páginaEl Abrazo de OsoMaximiliano ImazAún no hay calificaciones
- LISTA #1 Edf. HabitacionalesDocumento25 páginasLISTA #1 Edf. HabitacionalesKarlos MeihoffAún no hay calificaciones
- Etiqueta y Protocolo Tarea 1Documento2 páginasEtiqueta y Protocolo Tarea 1Leanny MarielAún no hay calificaciones
- 1 PresentaciónDocumento4 páginas1 PresentaciónPriscilla DavisAún no hay calificaciones
- Articulo Hijos Con Padres PresosDocumento4 páginasArticulo Hijos Con Padres PresosBeltrán Seco VillalobosAún no hay calificaciones
- Tarea 2 Empresa 1Documento17 páginasTarea 2 Empresa 1Marisol TomásAún no hay calificaciones
- Normas Peruanas de CementoDocumento1093 páginasNormas Peruanas de CementoAngel Washington Ochoa GalindoAún no hay calificaciones
- PlanesdecuidadopediatriaDocumento3 páginasPlanesdecuidadopediatriaAlejandra TorresAún no hay calificaciones
- 2 Sistemas de Propulsion de Cohetes PDFDocumento73 páginas2 Sistemas de Propulsion de Cohetes PDFTiare AntunezAún no hay calificaciones
- Lagunas - Milton LäuferDocumento165 páginasLagunas - Milton LäuferAlfredo BalanescuAún no hay calificaciones
- Jordi Barbeta. Ni Dios Ni PatrónDocumento1 páginaJordi Barbeta. Ni Dios Ni PatrónferrancabAún no hay calificaciones
- Mobiliario de Oficinas - VLK ProyectosDocumento27 páginasMobiliario de Oficinas - VLK ProyectosMiguel LimaAún no hay calificaciones
- Caso Idl3 - Informe Final Gestion de ProductoDocumento26 páginasCaso Idl3 - Informe Final Gestion de ProductoCarlos Crisostomo Cardenas50% (2)
- Carencias Nutricionales en Cítricos - Síntomas y CorrecciónDocumento9 páginasCarencias Nutricionales en Cítricos - Síntomas y CorrecciónslugmotionAún no hay calificaciones
- Aikido Tecnicas de Defensa Personal - PDFDocumento123 páginasAikido Tecnicas de Defensa Personal - PDFFederico Sebastian Carluccio100% (1)
- Cronograma de Seguridad Del PacienteDocumento4 páginasCronograma de Seguridad Del PacienteMaritza CasallasAún no hay calificaciones
- Derrame de Mercurio - ChoropampaDocumento9 páginasDerrame de Mercurio - ChoropampaHans BottgerAún no hay calificaciones
- AtletismoDocumento25 páginasAtletismoserviciosimedyilAún no hay calificaciones