También podría gustarte
- El Sonido, Música, Ingenieria Del SonidoDocumento34 páginasEl Sonido, Música, Ingenieria Del SonidoGarcía Frank100% (1)
- El Sonido Musica Ingenieria Del SonidoDocumento34 páginasEl Sonido Musica Ingenieria Del SonidoRicardo Ruano MartínezAún no hay calificaciones
- Ruido y VibracionesDocumento20 páginasRuido y VibracionesAbel BrunoriAún no hay calificaciones
- PSICOACÚSTICADocumento6 páginasPSICOACÚSTICAEuder Gómez VegaAún no hay calificaciones
- Sesión 1.1-La Física Del Sonido PDFDocumento10 páginasSesión 1.1-La Física Del Sonido PDFGonzalo RobledoAún no hay calificaciones
- El Origen Del Sonido y Sus PropiedadesDocumento6 páginasEl Origen Del Sonido y Sus PropiedadesPaul JacksonAún no hay calificaciones
- Fisiología de La AudiciónDocumento10 páginasFisiología de La AudiciónjurevecAún no hay calificaciones
- Qué Es El SonidoDocumento9 páginasQué Es El SonidoFidel LaimeAún no hay calificaciones
- Fundamento de Sonido y AudioDocumento11 páginasFundamento de Sonido y AudioJack GutiérrezAún no hay calificaciones
- Sonido y AcusticaDocumento20 páginasSonido y Acusticaapi-19885826Aún no hay calificaciones
- UD1 La Señal de AudioDocumento15 páginasUD1 La Señal de AudioFranciscoMolinaSánchezAún no hay calificaciones
- AcusticaDocumento15 páginasAcusticaLUVIANO PARTIDA JESUS ENRIQUEAún no hay calificaciones
- Acústica, SonidoDocumento55 páginasAcústica, SonidoEmilio ÁlvarezAún no hay calificaciones
- Parametros Subjetivos Del SonidoDocumento4 páginasParametros Subjetivos Del SonidoMartin CruzAún no hay calificaciones
- Voz Humana y Psicofísica de La AudiciónDocumento8 páginasVoz Humana y Psicofísica de La AudiciónCésar Arturo Aguilar BocanegraAún no hay calificaciones
- Intensidad SonoraDocumento28 páginasIntensidad SonoraCarol GiefAún no hay calificaciones
- Ruido y VibracionesDocumento51 páginasRuido y VibracionesAlejandraVilchesCastroAún no hay calificaciones
- Manual de Creación de Música ElectrónicaDocumento19 páginasManual de Creación de Música ElectrónicaAlberto Marcos Cabero100% (5)
- Informe - Manejo Del SonidoDocumento26 páginasInforme - Manejo Del SonidoErika VintimillaAún no hay calificaciones
- 3-1 - El Sonido Conceptos BsicosDocumento9 páginas3-1 - El Sonido Conceptos BsicosEleazar Moreno OrtizAún no hay calificaciones
- Clase de Sonido 1Documento9 páginasClase de Sonido 1JoeAún no hay calificaciones
- Tema 3 PERCEPCIÓN DE LA SONORIDAD Y LA DIRECCIONALIDADDocumento12 páginasTema 3 PERCEPCIÓN DE LA SONORIDAD Y LA DIRECCIONALIDADx3mgzAún no hay calificaciones
- AMPLITUDDocumento5 páginasAMPLITUDCamila AlvarezAún no hay calificaciones
- Intensidad de SonidoDocumento4 páginasIntensidad de Sonido조엘봉봉Aún no hay calificaciones
- SonidoDocumento2 páginasSonidoJairo CaballeroAún no hay calificaciones
- Lectura 13Documento6 páginasLectura 13Denis Peralta VillalobosAún no hay calificaciones
- Onda SonoraDocumento7 páginasOnda SonoraAna Sol Munizaga SaldañoAún no hay calificaciones
- Presentacion 9Documento67 páginasPresentacion 9kikeAún no hay calificaciones
- Clase 1 Leyes Del SonidoDocumento24 páginasClase 1 Leyes Del SonidoJacqueline Z. IzurragaAún no hay calificaciones
- La Percepción AuditivaDocumento23 páginasLa Percepción AuditivaRosa BañulsAún no hay calificaciones
- Capítulo 2. El RuidoDocumento56 páginasCapítulo 2. El Ruidohector hurtadoAún no hay calificaciones
- La Senal de Audio PDFDocumento26 páginasLa Senal de Audio PDFCarlos gomariZAún no hay calificaciones
- Fisica Del SonidoDocumento17 páginasFisica Del Sonidoulibarri83Aún no hay calificaciones
- Sonido GiancoliDocumento7 páginasSonido GiancoliPatricio Xavier Trujillo CaizaguanoAún no hay calificaciones
- Tema 9Documento16 páginasTema 9kikeAún no hay calificaciones
- Resumen Capítulo 1 y 2 MiyaraDocumento9 páginasResumen Capítulo 1 y 2 MiyaraOtonielAún no hay calificaciones
- Ondas SonorasDocumento26 páginasOndas SonorasNelson ChiribogaAún no hay calificaciones
- Curso Introductorio A La Sensibilización Sonora - Módulo IV & VDocumento11 páginasCurso Introductorio A La Sensibilización Sonora - Módulo IV & VAprendizReikiAún no hay calificaciones
- Seminario de Física Sesión 13 - TareaDocumento10 páginasSeminario de Física Sesión 13 - TareaDevora GilAún no hay calificaciones
- Fisica AcusticaDocumento7 páginasFisica AcusticaMauricio LustigAún no hay calificaciones
- SONIDO Guia de Estudios Primer Cuatrimestre 2022Documento85 páginasSONIDO Guia de Estudios Primer Cuatrimestre 2022Rama BieraAún no hay calificaciones
- Biofísica de La Audición y de La VisiónDocumento36 páginasBiofísica de La Audición y de La VisiónmikimedicinanAún no hay calificaciones
- Fisica de La AudicionDocumento5 páginasFisica de La AudicionKarlaPatriciaAún no hay calificaciones
- Física de La AudiciónDocumento19 páginasFísica de La AudiciónKaterin Hilario100% (2)
- Sonido 1Documento8 páginasSonido 1Alexis Exequiel RuppelAún no hay calificaciones
- Antologia Lenguaje SonoroDocumento28 páginasAntologia Lenguaje SonoroWilly Oliver PonceAún no hay calificaciones
- Session 11 - Environmental NoiseDocumento75 páginasSession 11 - Environmental NoiseMARIBEL EGUIZABAL QUISPEAún no hay calificaciones
- Qué Es El Sonido y Sus CaracteristicasDocumento2 páginasQué Es El Sonido y Sus CaracteristicasYermi HerreraAún no hay calificaciones
- 1resumen Fisica AcusticaDocumento8 páginas1resumen Fisica AcusticakamilaAún no hay calificaciones
- PDF Esp 382 PDFDocumento21 páginasPDF Esp 382 PDFRubenAún no hay calificaciones
- Ruido TPDocumento22 páginasRuido TPMaria Eugenia Ferro100% (1)
- Fonética y Fonología Tema 3Documento10 páginasFonética y Fonología Tema 3AndreaAún no hay calificaciones
- Apunte Telco 1er Parcial Wiernik Con Respuestas Kahoot-ComprimidoDocumento61 páginasApunte Telco 1er Parcial Wiernik Con Respuestas Kahoot-ComprimidoMariano Rubén JurczyszynAún no hay calificaciones
- Resumen El SonidoDocumento14 páginasResumen El Sonidocesar marconiAún no hay calificaciones
- Guia8a SonidoYSensibilidadAuditiva TeoriaDocumento6 páginasGuia8a SonidoYSensibilidadAuditiva TeoriaJean Franco OquendoAún no hay calificaciones
- Apuntes de Clases - Todos-Insta IiDocumento24 páginasApuntes de Clases - Todos-Insta IiClaudia CoronelAún no hay calificaciones
- Ecografía normal del árbol urinario y genitales externosDe EverandEcografía normal del árbol urinario y genitales externosAún no hay calificaciones
- La sanación a través del sonido para principiantes: Medicina sónica para el cuerpo, rituales de chakras y lo que no te han contado sobre la energía vibracionalDe EverandLa sanación a través del sonido para principiantes: Medicina sónica para el cuerpo, rituales de chakras y lo que no te han contado sobre la energía vibracionalAún no hay calificaciones
- Descontruccion IdeologiacaDocumento1 páginaDescontruccion IdeologiacaUriel Antonio Rodríguez VillarrealAún no hay calificaciones
- Lesiones Benignas de La LaringeDocumento18 páginasLesiones Benignas de La LaringeUriel Antonio Rodríguez VillarrealAún no hay calificaciones
- Casos ClinicosDocumento5 páginasCasos ClinicosUriel Antonio Rodríguez VillarrealAún no hay calificaciones
- Tumor de OrofaringeDocumento16 páginasTumor de OrofaringeUriel Antonio Rodríguez VillarrealAún no hay calificaciones
- Apnea Del Sueño y RonquidosDocumento8 páginasApnea Del Sueño y RonquidosUriel Antonio Rodríguez VillarrealAún no hay calificaciones
- Anatomia, Embriologia y FisiologiaDocumento4 páginasAnatomia, Embriologia y FisiologiaUriel Antonio Rodríguez VillarrealAún no hay calificaciones
- Eckert FisioAnimal PDFDocumento694 páginasEckert FisioAnimal PDFArturo AirgunAún no hay calificaciones
- Sistema NerviosoDocumento126 páginasSistema NerviosoDaniel Flores PérezAún no hay calificaciones
- Resúmen de Capítulos de Dragones Del EdénDocumento5 páginasResúmen de Capítulos de Dragones Del EdénLucia Tello Moscarella50% (2)
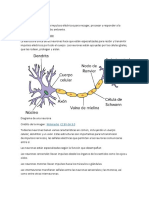
- Propiedades de La NeuronaDocumento2 páginasPropiedades de La NeuronaDimasramirezAún no hay calificaciones
- Bases Físicas de Los Fenómenos BioeléctricosDocumento65 páginasBases Físicas de Los Fenómenos BioeléctricosAndrew Emilio Castillo Pacheco0% (1)
- Resumen - El Cerebro y La ConductaDocumento3 páginasResumen - El Cerebro y La ConductaAkary Jocelyn Ruiz100% (1)
- 05 Sist - Estomatognatico Recept. RefDocumento37 páginas05 Sist - Estomatognatico Recept. RefVladimir Sánchez Chávez-ArroyoAún no hay calificaciones
- Sistema Nervioso 01Documento10 páginasSistema Nervioso 01Nancy Yadira Rodriguez GalvisAún no hay calificaciones
- El Sistema NerviosoDocumento3 páginasEl Sistema NerviosoIsa SantiagoAún no hay calificaciones
- La Quimica de La FelicidadDocumento12 páginasLa Quimica de La FelicidadJuan José V. CáceresAún no hay calificaciones
- Las Ecuaciones Del Libre AlbedríoDocumento19 páginasLas Ecuaciones Del Libre AlbedríoIbrah GabriellAún no hay calificaciones
- Resumen: El Sistema NerviosoDocumento9 páginasResumen: El Sistema NerviosoISABELLE GONZaLEZAún no hay calificaciones
- Neurodesarrollo Infantil (Orlando Torre María Priscilla)Documento219 páginasNeurodesarrollo Infantil (Orlando Torre María Priscilla)Rey Vir100% (1)
- Algodistrofia o Sindrome Doloroso Regional Compejo Tipo 1Documento12 páginasAlgodistrofia o Sindrome Doloroso Regional Compejo Tipo 1Ivan Hernandez Hernandez100% (2)
- 8° Guias 3 Periodo 2 Sistemas Nervioso, Receptores SensorialesDocumento5 páginas8° Guias 3 Periodo 2 Sistemas Nervioso, Receptores SensorialesMari Fernanda Méndez OchoaAún no hay calificaciones
- Que Hago Si Mi Media Naranja Es ToronjaDocumento11 páginasQue Hago Si Mi Media Naranja Es ToronjaNalu Muñoz0% (1)
- Resumen Completo Todo FarmaDocumento213 páginasResumen Completo Todo FarmaRomario CamachoAún no hay calificaciones
- Inteligencia InformeDocumento48 páginasInteligencia InformeJose Cueva ArevaloAún no hay calificaciones
- Los Trastornos Del - Espectro AutistaDocumento202 páginasLos Trastornos Del - Espectro AutistaMiguel AtemporaliaAún no hay calificaciones
- Metodo de KaniDocumento13 páginasMetodo de KaniTejerina SFAún no hay calificaciones
- Echeverría Meily U1 Informe2Documento3 páginasEcheverría Meily U1 Informe2Meily BorjasAún no hay calificaciones
- Psicologia CepruDocumento83 páginasPsicologia CepruRamos Ventura HenryAún no hay calificaciones
- CHERTOK Las Causas de Nuestra ConductaDocumento9 páginasCHERTOK Las Causas de Nuestra ConductailanaAún no hay calificaciones
- La Luz Del AlmaDocumento49 páginasLa Luz Del AlmaJoseAún no hay calificaciones
- Conceptos Del Condicionamiento Clásico en Los Campos Básicos y Aplicados de La PsicologíaDocumento15 páginasConceptos Del Condicionamiento Clásico en Los Campos Básicos y Aplicados de La Psicologíaanahata.conectadosconelcorazonAún no hay calificaciones
- Resumen U 1,2,3,4,5 Neuro 1Documento31 páginasResumen U 1,2,3,4,5 Neuro 1IaraCamila GuerraAún no hay calificaciones
- El Arpa Mágica - Metafísica para TodosDocumento79 páginasEl Arpa Mágica - Metafísica para TodosIzayshebas Corp100% (1)
- Sistema NerviosoDocumento5 páginasSistema NerviosoDavid QuinteroAún no hay calificaciones
- Hoja de TrabajoDocumento5 páginasHoja de TrabajoLeandro E D U A R D OAún no hay calificaciones
- Inteligencia ProspectivaDocumento130 páginasInteligencia Prospectivanelson bary serna marcos100% (4)