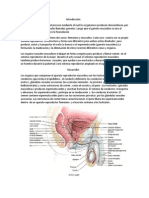
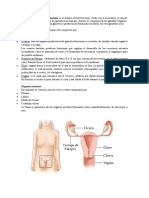
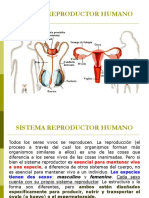
También podría gustarte
- Sistema Reproductor MasculinoDocumento7 páginasSistema Reproductor Masculinoconrado GuillermoAún no hay calificaciones
- Apuntes Anatomía (24 Pag.) .Documento24 páginasApuntes Anatomía (24 Pag.) .JosefaAún no hay calificaciones
- Af1-Mod 8-ReprDocumento31 páginasAf1-Mod 8-ReprGustavo CamaraAún no hay calificaciones
- Material Bibliografico de Biologia-3 Año-23Documento10 páginasMaterial Bibliografico de Biologia-3 Año-23betianaolmedo2023Aún no hay calificaciones
- Reproducción humanaDocumento11 páginasReproducción humanaMeybalex GAAún no hay calificaciones
- Taller de Reproducción HumanaDocumento24 páginasTaller de Reproducción HumanaMaryoris Del C. Osorio DE LA OssaAún no hay calificaciones
- Reproducción Sexual en Humanos. Parte 1Documento6 páginasReproducción Sexual en Humanos. Parte 1pokeraperoAún no hay calificaciones
- Aparatos ReproductoresDocumento38 páginasAparatos ReproductoresAlisson CahuanaAún no hay calificaciones
- Aparato reproductor masculino y femeninoDocumento3 páginasAparato reproductor masculino y femeninoYaira Nicolle Cujia NievesAún no hay calificaciones
- UntitledDocumento19 páginasUntitledNajara HookerAún no hay calificaciones
- Aparato reproductor humano: órganos y procesosDocumento26 páginasAparato reproductor humano: órganos y procesosargaxdAún no hay calificaciones
- El Aparato Reproductor MasculinoDocumento13 páginasEl Aparato Reproductor MasculinoArgenisAún no hay calificaciones
- AP - Reproductor MasculinoDocumento12 páginasAP - Reproductor MasculinoantonellamartAún no hay calificaciones
- Aparato Reproductor Femenino y MasculinoDocumento22 páginasAparato Reproductor Femenino y MasculinoAngela ChengAún no hay calificaciones
- Tema 17 CCNN 1ro Sec Sistema Reproductor HumanoDocumento4 páginasTema 17 CCNN 1ro Sec Sistema Reproductor HumanoLuis Guillermo Pareja PevesAún no hay calificaciones
- Reproducción en El MachoDocumento7 páginasReproducción en El Machoestudiante veterinarioAún no hay calificaciones
- Sistema Reproductor Masculino. InformeDocumento10 páginasSistema Reproductor Masculino. Informemanuel quintanillaAún no hay calificaciones
- PRÁCTICA No. 9Documento17 páginasPRÁCTICA No. 9Génesis RomeroAún no hay calificaciones
- Módulo 7 Med Ind 2016Documento10 páginasMódulo 7 Med Ind 2016Eliana VelozoAún no hay calificaciones
- Sistema reproductor animales producciónDocumento12 páginasSistema reproductor animales producciónAlvaroJJaimesCAún no hay calificaciones
- ReproduccionDocumento28 páginasReproduccionLuis Xavier Sanchez S.Aún no hay calificaciones
- Repaso Estructura NO.3Documento8 páginasRepaso Estructura NO.3Anthony AntunaAún no hay calificaciones
- Sistema Reproductor MasculinoDocumento6 páginasSistema Reproductor Masculinocheilo eren doxAún no hay calificaciones
- BIOLOGIA UNIDAD 3Documento16 páginasBIOLOGIA UNIDAD 3francolopez327Aún no hay calificaciones
- Aparato Reproductor de Ambos Sexos.Documento8 páginasAparato Reproductor de Ambos Sexos.Tanisha Mercado PeñaAún no hay calificaciones
- Sistema Reproductor. ApuntesDocumento6 páginasSistema Reproductor. ApuntesRubénDaríoAún no hay calificaciones
- Sistema reproductor masculinoDocumento12 páginasSistema reproductor masculinoNancydeValenciaAún no hay calificaciones
- Aparato Reproductor Femenino y MasculinoDocumento10 páginasAparato Reproductor Femenino y MasculinojeanniaAún no hay calificaciones
- Sistema Reproductor Masculino W 1Documento31 páginasSistema Reproductor Masculino W 1ER Tatiana RiosAún no hay calificaciones
- Tema - SISTEMA REPRODUCTOR HUMANODocumento24 páginasTema - SISTEMA REPRODUCTOR HUMANOLupita LeonardoAún no hay calificaciones
- TP 2 - Aparato ReporductorDocumento5 páginasTP 2 - Aparato Reporductormagali.valcarcelAún no hay calificaciones
- Módulo 2Documento49 páginasMódulo 2pilar valenciaAún no hay calificaciones
- Biología Humana Resumen 1Documento34 páginasBiología Humana Resumen 1Luz MugnaAún no hay calificaciones
- Andrologia Clase 1Documento28 páginasAndrologia Clase 1Anthony VegaAún no hay calificaciones
- Informe ReproduccionDocumento8 páginasInforme ReproduccionLucia Amanda AuroraAún no hay calificaciones
- Documento Sin TítuloDocumento5 páginasDocumento Sin TítuloLoreanney FrancoAún no hay calificaciones
- Guido Beitia SaludDocumento8 páginasGuido Beitia SaludFranco BaldaniAún no hay calificaciones
- El Sistema Reproductor Humano PDFDocumento33 páginasEl Sistema Reproductor Humano PDFedwinAún no hay calificaciones
- Tema 7 - La ReproducciónDocumento11 páginasTema 7 - La Reproducciónmaytemasa100% (1)
- Tarea Ciencias Sis Rep y Sis NerviosoDocumento9 páginasTarea Ciencias Sis Rep y Sis Nerviosopaulafer2014Aún no hay calificaciones
- Anatomía y fisiología del aparato reproductorDocumento12 páginasAnatomía y fisiología del aparato reproductormercedes martinez de toroAún no hay calificaciones
- Proyecto de FisiologiaDocumento21 páginasProyecto de FisiologiaCruz Noemi Orozco TerwogtAún no hay calificaciones
- Sistema Reproductor - MonografiaDocumento38 páginasSistema Reproductor - MonografiaDaniel Rivera SilvestreAún no hay calificaciones
- Anatomía y Fisiología Del Aparato Reproductor Masculino y FemeninoDocumento53 páginasAnatomía y Fisiología Del Aparato Reproductor Masculino y FemeninoReyes Salazar100% (9)
- Sistema Reproductor Masculino-FabioDocumento11 páginasSistema Reproductor Masculino-FabioFabio ZarateAún no hay calificaciones
- Órganos Sexuales PDFDocumento20 páginasÓrganos Sexuales PDFPerez Tuxpan Jesus AlejandroAún no hay calificaciones
- Reproducción humana: Gametos, fecundación y desarrollo embrionarioDocumento9 páginasReproducción humana: Gametos, fecundación y desarrollo embrionarioIngrid Tatiana HolguinAún no hay calificaciones
- Sistema reproductor cerdoDocumento7 páginasSistema reproductor cerdoUziel Rojas100% (1)
- Aparato ReproductorDocumento26 páginasAparato ReproductorAirinth Rodriguez GiraldoAún no hay calificaciones
- Estructuras Del Sistema Reproductivo y Sus FuncionesDocumento8 páginasEstructuras Del Sistema Reproductivo y Sus FuncionesSebastian Vega HernandezAún no hay calificaciones
- Aparato Reproductor Masculino TejidosDocumento11 páginasAparato Reproductor Masculino Tejidospanther100Aún no hay calificaciones
- Aparato Reproductor Femenino y MasculinoDocumento10 páginasAparato Reproductor Femenino y MasculinoDenisse ValleAún no hay calificaciones
- Aparato Reproductor MasculinoDocumento11 páginasAparato Reproductor Masculinoricary rodriguezAún no hay calificaciones
- Aparato Reproductor Femenino InformeDocumento7 páginasAparato Reproductor Femenino InformeHans JuanAún no hay calificaciones
- FisiologíaDocumento15 páginasFisiologíaBell GameroAún no hay calificaciones
- Aparato Reproductor MasculinoDocumento6 páginasAparato Reproductor MasculinoLuis FernandoAún no hay calificaciones
- Aparato Reproductor MasculinoDocumento5 páginasAparato Reproductor MasculinoCOMERCIAL GARCIAAún no hay calificaciones
- Aparato Reproductor Masculino Final HOMBREDocumento8 páginasAparato Reproductor Masculino Final HOMBREMauricio SanchezAún no hay calificaciones
- 7 Aparato ReproductorDocumento24 páginas7 Aparato ReproductorCuba CcAún no hay calificaciones
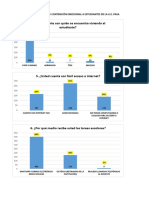
- Tabulación Encuesta GeneralDocumento4 páginasTabulación Encuesta GeneralAlexLlumiquingaAún no hay calificaciones
- Lugo Luis Collage Conceptos en Salud Pública en El HogarDocumento6 páginasLugo Luis Collage Conceptos en Salud Pública en El Hogardiego baronAún no hay calificaciones
- Composición Química de La Gastrina y Otras HormonasDocumento16 páginasComposición Química de La Gastrina y Otras HormonasHeidi PerezAún no hay calificaciones
- Neuropsicología de La AdicciónDocumento54 páginasNeuropsicología de La AdicciónDavid CarrilloAún no hay calificaciones
- Perfiles AnaliticosDocumento12 páginasPerfiles AnaliticosUnidad MolecularAún no hay calificaciones
- Programa II Jornadas ViEDocumento36 páginasPrograma II Jornadas ViEDiana FlaquéAún no hay calificaciones
- 2.vias de Administracion de FarmacosDocumento28 páginas2.vias de Administracion de FarmacosLizeth Margarita Cassiani HerreraAún no hay calificaciones
- Guia de Higiene (1) - TaggedDocumento19 páginasGuia de Higiene (1) - TaggedMIRELLA ABIGAIL LLOCLLA MOROCHOAún no hay calificaciones
- Carla Andrea Camacho Villalobos 2018 PDFDocumento9 páginasCarla Andrea Camacho Villalobos 2018 PDFCarlaAndreaCamachoVillalobosAún no hay calificaciones
- Ficha Cosecha de Uva en Viña ODIDocumento2 páginasFicha Cosecha de Uva en Viña ODIJuan Pablo Moya AzocarAún no hay calificaciones
- Defensa ProyectoDocumento4 páginasDefensa ProyectoIsabel Heras CortésAún no hay calificaciones
- Historia de La MedicinaDocumento14 páginasHistoria de La MedicinafiorellaAún no hay calificaciones
- Climatologia MedicaDocumento10 páginasClimatologia MedicaMaría Arahuallpa100% (1)
- S.pers.04 NovDocumento5 páginasS.pers.04 NovLIBRERIA EL AMIGOAún no hay calificaciones
- Fascículo UNIDAD 3 - Curso Competencias Socioemocionales PDFDocumento26 páginasFascículo UNIDAD 3 - Curso Competencias Socioemocionales PDFDavid Cabana HuanquiAún no hay calificaciones
- Los Engramas y Nuestros Universos PsíquicosDocumento2 páginasLos Engramas y Nuestros Universos PsíquicosIreneAún no hay calificaciones
- Capacitación de Seguridad y Salud en El Trabajo BPMDocumento14 páginasCapacitación de Seguridad y Salud en El Trabajo BPMJulian Andres Salamanca OrozcoAún no hay calificaciones
- Formatos Oe Lurawi Peru 2022 OkDocumento18 páginasFormatos Oe Lurawi Peru 2022 OkBessy HuamanAún no hay calificaciones
- Información estudianteDocumento7 páginasInformación estudianteYenny FernandezAún no hay calificaciones
- Árbol de ProblemasDocumento2 páginasÁrbol de ProblemasJuana Mejia SaavedraAún no hay calificaciones
- Act 2 Tecnicas Proyectivas 1Documento6 páginasAct 2 Tecnicas Proyectivas 1Dan Isai Rosales100% (1)
- T.P 1 HistoriaDocumento6 páginasT.P 1 HistoriaPameAún no hay calificaciones
- Estres Laboral y Pausas ActivasDocumento50 páginasEstres Laboral y Pausas ActivasTony C CampAún no hay calificaciones
- Previser Folleto BienestarDocumento68 páginasPreviser Folleto BienestarELmasterAún no hay calificaciones
- Tendencias Actuales en La Planificación Del EntrenamientoDocumento7 páginasTendencias Actuales en La Planificación Del EntrenamientoMaikel GarciaAún no hay calificaciones
- Eval - Diagnostico L.L.Documento12 páginasEval - Diagnostico L.L.Ele AyoviAún no hay calificaciones
- Trastornos de la motricidad orofacial en niños: causas, clasificación y manejoDocumento6 páginasTrastornos de la motricidad orofacial en niños: causas, clasificación y manejoJaviera Guzmán SoAún no hay calificaciones
- Glosario Cuaderno de Neurociencia NetterDocumento84 páginasGlosario Cuaderno de Neurociencia NetterRosana Edith BravoAún no hay calificaciones
- Técnica de Abordaje Del Estudio Suprapúbico y Transvaginal en GinecobstetriciaDocumento6 páginasTécnica de Abordaje Del Estudio Suprapúbico y Transvaginal en GinecobstetriciaIris De Los SantosAún no hay calificaciones
- SEMINARIODocumento9 páginasSEMINARIOZaray EspinosaAún no hay calificaciones