También podría gustarte
- FICHA TECNICA - TuboPlusDocumento6 páginasFICHA TECNICA - TuboPlusMiguel AngelAún no hay calificaciones
- Sea Psiquico FinalDocumento241 páginasSea Psiquico FinalFernando Javier LeivaAún no hay calificaciones
- Tablas de ElectricidadDocumento6 páginasTablas de ElectricidadLeandro HerreraAún no hay calificaciones
- Chancado de MineralesDocumento49 páginasChancado de MineralesRAFAEL CHAVEZ YAURIAún no hay calificaciones
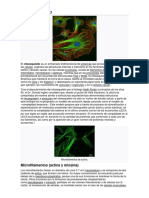
- Citoesqueleto y Motilidad Celular. Filamentos 14-07-2016Documento63 páginasCitoesqueleto y Motilidad Celular. Filamentos 14-07-2016Beto BarcelonaAún no hay calificaciones
- Flotación cal vs pHDocumento16 páginasFlotación cal vs pHValentinaAún no hay calificaciones
- CitoesqueletoDocumento11 páginasCitoesqueletoGabriela PeñaAún no hay calificaciones
- CitoesqueletoDocumento21 páginasCitoesqueletoSoraida Sanchez ParraAún no hay calificaciones
- Citoesqueleto OriginalDocumento16 páginasCitoesqueleto OriginalfranciscoAún no hay calificaciones
- CitoesqueletoDocumento11 páginasCitoesqueletoMilagros QuispeAún no hay calificaciones
- El Citoesquelet1Documento6 páginasEl Citoesquelet1sandraAún no hay calificaciones
- Cito EsqueletoDocumento4 páginasCito EsqueletoMateo DuraznoAún no hay calificaciones
- ENSAYO Citoesqueleto Karisbel SDocumento4 páginasENSAYO Citoesqueleto Karisbel Skarisbel suarezAún no hay calificaciones
- Citoesqueleto y MovimientoDocumento14 páginasCitoesqueleto y Movimientoandreaave100% (2)
- MICROTUBULOSDocumento4 páginasMICROTUBULOSdaniel feijooAún no hay calificaciones
- CITOESQUELETO GenomasurDocumento32 páginasCITOESQUELETO GenomasurDaiana tabordaAún no hay calificaciones
- Motilidad CelularDocumento4 páginasMotilidad CelularFrancis NicoleAún no hay calificaciones
- CitoesqueletoDocumento9 páginasCitoesqueletoOscar RomeroAún no hay calificaciones
- Introducción Al CitoesqueletoDocumento5 páginasIntroducción Al CitoesqueletoAina cuixart gimoAún no hay calificaciones
- Que Son Los Microtubulos Filamentos IntermediosDocumento6 páginasQue Son Los Microtubulos Filamentos IntermediosMarvinRiveraAún no hay calificaciones
- CitoesqueletoDocumento8 páginasCitoesqueletoMili SassoneAún no hay calificaciones
- Estructuras y Organulos No MebranososDocumento7 páginasEstructuras y Organulos No MebranososRaul Ibanez RodriguezAún no hay calificaciones
- El Citoesqueleto Es Una Estructura Dinámica de Las Células Eucariotas Que Permite Mantener o Cambiar La Forma Celular Reaccionando A Estímulos Externos o InternosDocumento2 páginasEl Citoesqueleto Es Una Estructura Dinámica de Las Células Eucariotas Que Permite Mantener o Cambiar La Forma Celular Reaccionando A Estímulos Externos o InternosmitzyjackyAún no hay calificaciones
- Unidad 9 CitoesqueletoDocumento43 páginasUnidad 9 CitoesqueletoAlba Fernández RevueltaAún no hay calificaciones
- CitoesqueletoDocumento8 páginasCitoesqueletoAlejandra GuaillasAún no hay calificaciones
- 05 CITOPLASMA - CITOESQUELETODocumento19 páginas05 CITOPLASMA - CITOESQUELETOAlfredo Sam Alvaro HerreraAún no hay calificaciones
- Estructura y funciones del citoesqueletoDocumento32 páginasEstructura y funciones del citoesqueletoGabriela IbañezAún no hay calificaciones
- Citoesqueleto PresentacionDocumento13 páginasCitoesqueleto PresentacionRachel CardenasAún no hay calificaciones
- Citoesqueleto PresentaciónDocumento15 páginasCitoesqueleto PresentaciónLauAún no hay calificaciones
- EL CITOesqueletoDocumento4 páginasEL CITOesqueletoDiana Del Pilar Chaustre DuranAún no hay calificaciones
- Cito EsqueletoDocumento8 páginasCito EsqueletoJuan OrtízAún no hay calificaciones
- Citoesqueleto: FuncionesDocumento14 páginasCitoesqueleto: Funcioneskarina ramos100% (1)
- 6 Ensayo CitoesqueletoDocumento6 páginas6 Ensayo CitoesqueletoValeria Fernanda Rocha MerinoAún no hay calificaciones
- CITOESQUELETO TextoDocumento9 páginasCITOESQUELETO Textoeven24Aún no hay calificaciones
- Forma y MovimientoDocumento12 páginasForma y MovimientoPichardo Franco Melanie VioletaAún no hay calificaciones
- Citoesqueleto Seminario 2Documento18 páginasCitoesqueleto Seminario 2Luu GonzalezAún no hay calificaciones
- Citoesqueleto y ciclo celularDocumento15 páginasCitoesqueleto y ciclo celularDanna Valentina MartinezAún no hay calificaciones
- CitoesqueletoDocumento13 páginasCitoesqueletoBian RamirezAún no hay calificaciones
- CITOESQUELETODocumento29 páginasCITOESQUELETOElisandra Rodriguez GonzalesAún no hay calificaciones
- Citoesqueleto 2021Documento70 páginasCitoesqueleto 2021Fran TejadaAún no hay calificaciones
- Cito EsqueletoDocumento8 páginasCito EsqueletoMarilyn MontecinosAún no hay calificaciones
- CitoesqueletoDocumento102 páginasCitoesqueletoMarco David Meza SerpaAún no hay calificaciones
- Biología P19Documento15 páginasBiología P19jg2488955Aún no hay calificaciones
- El CitoesqueletoDocumento7 páginasEl CitoesqueletoKateryn ÁlvarezAún no hay calificaciones
- TripticoDocumento9 páginasTripticojesus jimenezAún no hay calificaciones
- Citoesqueleto 1Documento18 páginasCitoesqueleto 1Santiago PalaciosAún no hay calificaciones
- UNIDAD 5 CitoesqueletoDocumento58 páginasUNIDAD 5 CitoesqueletoCarolina Bravo BastidasAún no hay calificaciones
- Organelos de La CélulaDocumento16 páginasOrganelos de La CélulaSu SaucedoAún no hay calificaciones
- Guía de Diapositivas MaferDocumento29 páginasGuía de Diapositivas MaferGuidmari ValeroAún no hay calificaciones
- Citoesqueleto 2023Documento68 páginasCitoesqueleto 2023FlorAún no hay calificaciones
- Centriolos y citoesqueletoDocumento17 páginasCentriolos y citoesqueletovalentina DominguezAún no hay calificaciones
- Citoesqueleto Final IntroDocumento6 páginasCitoesqueleto Final IntroBelenMinAún no hay calificaciones
- Citoesqueleto: Estructura y funciones clave de los microfilamentos, filamentos intermedios y microtúbulosDocumento5 páginasCitoesqueleto: Estructura y funciones clave de los microfilamentos, filamentos intermedios y microtúbulosErika Maribel BadilloAún no hay calificaciones
- Tarea 14Documento8 páginasTarea 14Paula LopezAún no hay calificaciones
- CITOESQUELETODocumento26 páginasCITOESQUELETOMinerva ShapplinAún no hay calificaciones
- El citoesqueleto: microtúbulos, funciones y estructuraDocumento9 páginasEl citoesqueleto: microtúbulos, funciones y estructuraMercedes MartínAún no hay calificaciones
- Sesión 3 Teoría Celular. Célula Procariótica y Eucariótica. CitosolDocumento55 páginasSesión 3 Teoría Celular. Célula Procariótica y Eucariótica. CitosolLEYDI ARIANA CRUZ GARCIAAún no hay calificaciones
- Citoplasma, Citoesqueleto, RibosomasDocumento26 páginasCitoplasma, Citoesqueleto, Ribosomas2cr_28Aún no hay calificaciones
- CitoesqueletoDocumento3 páginasCitoesqueletoJosué SaavAún no hay calificaciones
- El citoesqueleto: estructura, funciones y componentesDocumento6 páginasEl citoesqueleto: estructura, funciones y componentesevelyn obregon jimenezAún no hay calificaciones
- Apunte CitoesqueletoDocumento4 páginasApunte CitoesqueletoAliciaAún no hay calificaciones
- Citoplasma y Citoesqueleto PDFDocumento19 páginasCitoplasma y Citoesqueleto PDFEver Villanueva100% (1)
- CitoesqueletoDocumento2 páginasCitoesqueletoLuis Tapia RubioAún no hay calificaciones
- P8. HPLCDocumento11 páginasP8. HPLCCarmen RodríguezAún no hay calificaciones
- La Estática en La Vida CotidianaDocumento315 páginasLa Estática en La Vida CotidianaDavidespin0% (1)
- CTA - U6 - 1er Grado - Sesion 03Documento5 páginasCTA - U6 - 1er Grado - Sesion 03yamiliAún no hay calificaciones
- Ley Inercia Ejemplos CotidianoDocumento9 páginasLey Inercia Ejemplos CotidianoJohana CubillosAún no hay calificaciones
- Razón de cambio promedio y razón de cambio instantáneoDocumento4 páginasRazón de cambio promedio y razón de cambio instantáneoLuis Alberto TRELLES ALVAREZAún no hay calificaciones
- Ciencia-Mostra-Primera-edicion-SoberaniaDocumento53 páginasCiencia-Mostra-Primera-edicion-SoberaniamagfvaldezAún no hay calificaciones
- Laboratorio Sobre Reflexión y RefracciónDocumento7 páginasLaboratorio Sobre Reflexión y Refracciónpedro pablo rosario vargasAún no hay calificaciones
- Campo Eléctrico 4to SecDocumento7 páginasCampo Eléctrico 4to SecChristianHansVillanuevaZuloetaAún no hay calificaciones
- Fundamentos de Maquinas ElectricasDocumento11 páginasFundamentos de Maquinas ElectricasIsaac GonzalezAún no hay calificaciones
- Contenido - Unidad 1Documento7 páginasContenido - Unidad 1Yilda Suero TejadaAún no hay calificaciones
- Ondas en Una Columna de AireDocumento6 páginasOndas en Una Columna de AireJsus Ramirez Jr.Aún no hay calificaciones
- Micrómetro y comparador de carátulaDocumento17 páginasMicrómetro y comparador de carátulagiovanny enrique altamarAún no hay calificaciones
- (H9) Respuestas Estatica 2004Documento10 páginas(H9) Respuestas Estatica 2004Imeldo GuerraAún no hay calificaciones
- Lista de Precios Inasel Siemens 2015pdfDocumento147 páginasLista de Precios Inasel Siemens 2015pdfKaryna GonzalezAún no hay calificaciones
- F0 - Sólidos GeométricosDocumento1 páginaF0 - Sólidos GeométricossurvivaldubcommcAún no hay calificaciones
- Unidad Didactica Energia GeotermicaDocumento24 páginasUnidad Didactica Energia GeotermicaAbel Damian OñaAún no hay calificaciones
- ReverberaciónDocumento17 páginasReverberaciónDiego MoraAún no hay calificaciones
- ICO-CAP1S2 Calculo Aplicado-U01 Geometria AnaliticaDocumento109 páginasICO-CAP1S2 Calculo Aplicado-U01 Geometria AnaliticaIgor DavalosAún no hay calificaciones
- Polietileno bimodal catalizadoresDocumento2 páginasPolietileno bimodal catalizadoresdianaAún no hay calificaciones
- CURVACEDocumento6 páginasCURVACESneider LandinezAún no hay calificaciones
- Consigna Examen Parcial GQT - Docx LaccDocumento8 páginasConsigna Examen Parcial GQT - Docx LaccJANET KARINA TORRES HUILLCAAún no hay calificaciones
- Evi2 CD LDGDocumento5 páginasEvi2 CD LDGluisAún no hay calificaciones
- Semana 9 y 10Documento15 páginasSemana 9 y 10Renato Rios RomeroAún no hay calificaciones
- MEMORIA DE CALCULO DE AsentamientosDocumento2 páginasMEMORIA DE CALCULO DE AsentamientosAlmendra FonsecaAún no hay calificaciones
- Guía PSU Cuerpos GeométricosDocumento4 páginasGuía PSU Cuerpos GeométricosVictor ParraAún no hay calificaciones