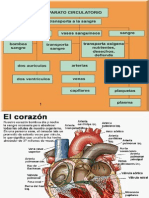
También podría gustarte
- CITOESQUELETODocumento29 páginasCITOESQUELETOElisandra Rodriguez GonzalesAún no hay calificaciones
- CitoesqueletoDocumento13 páginasCitoesqueletoBian RamirezAún no hay calificaciones
- Cito EsqueletoDocumento8 páginasCito EsqueletoMarilyn MontecinosAún no hay calificaciones
- CitoesqueletoDocumento21 páginasCitoesqueletoSoraida Sanchez ParraAún no hay calificaciones
- Citoesqueleto OriginalDocumento16 páginasCitoesqueleto OriginalfranciscoAún no hay calificaciones
- Citoplasma y citoesqueleto: componentes y funcionesDocumento50 páginasCitoplasma y citoesqueleto: componentes y funcionesyaritzaAún no hay calificaciones
- Citoesqueleto y MovimientoDocumento14 páginasCitoesqueleto y Movimientoandreaave100% (2)
- Citoesqueleto Final IntroDocumento6 páginasCitoesqueleto Final IntroBelenMinAún no hay calificaciones
- CitoesqueletoDocumento11 páginasCitoesqueletoMilagros QuispeAún no hay calificaciones
- CITOESQUELETODocumento23 páginasCITOESQUELETOMeritxell Planell CastellsAún no hay calificaciones
- Motilidad CelularDocumento4 páginasMotilidad CelularFrancis NicoleAún no hay calificaciones
- CitoesqueletoDocumento68 páginasCitoesqueletoGiovanna B86% (7)
- Citoesqueleto Lectura AdicionalDocumento3 páginasCitoesqueleto Lectura AdicionalNancy VielmanAún no hay calificaciones
- Citoesqueleto: Microfilamentos Filamentos Intermedios MicrotúbulosDocumento10 páginasCitoesqueleto: Microfilamentos Filamentos Intermedios MicrotúbulosDian GAún no hay calificaciones
- CITOESQUELETODocumento19 páginasCITOESQUELETOandrea rochaAún no hay calificaciones
- Estructura y funciones del citoesqueletoDocumento32 páginasEstructura y funciones del citoesqueletoGabriela IbañezAún no hay calificaciones
- Guía Trabajo Autónomo Citoesqueleto - ListaDocumento6 páginasGuía Trabajo Autónomo Citoesqueleto - ListaRoberto MarceliAún no hay calificaciones
- MICROTUBULOSDocumento4 páginasMICROTUBULOSdaniel feijooAún no hay calificaciones
- Citoesqueleto y ciclo celularDocumento15 páginasCitoesqueleto y ciclo celularDanna Valentina MartinezAún no hay calificaciones
- Sesión 3 Teoría Celular. Célula Procariótica y Eucariótica. CitosolDocumento55 páginasSesión 3 Teoría Celular. Célula Procariótica y Eucariótica. CitosolLEYDI ARIANA CRUZ GARCIAAún no hay calificaciones
- CitoesqueletoDocumento8 páginasCitoesqueletoMili SassoneAún no hay calificaciones
- CitoesqueletoDocumento6 páginasCitoesqueletodmejiagaAún no hay calificaciones
- Movimiento celular: Citoesqueleto y sus componentesDocumento28 páginasMovimiento celular: Citoesqueleto y sus componentesdante ulmer medina condorAún no hay calificaciones
- Citoplasma, Citoesqueleto, RibosomasDocumento26 páginasCitoplasma, Citoesqueleto, Ribosomas2cr_28Aún no hay calificaciones
- Cito EsqueletoDocumento8 páginasCito EsqueletoJuan OrtízAún no hay calificaciones
- Citoesqueleto PresentaciónDocumento15 páginasCitoesqueleto PresentaciónLauAún no hay calificaciones
- Unidad 9 CitoesqueletoDocumento43 páginasUnidad 9 CitoesqueletoAlba Fernández RevueltaAún no hay calificaciones
- CitoesqueletoDocumento9 páginasCitoesqueletoOscar RomeroAún no hay calificaciones
- ENSAYO Citoesqueleto Karisbel SDocumento4 páginasENSAYO Citoesqueleto Karisbel Skarisbel suarezAún no hay calificaciones
- HOja de Trabajo Semana 13 - 1Documento7 páginasHOja de Trabajo Semana 13 - 1Estrella PalaciosAún no hay calificaciones
- MicrotúbulosDocumento3 páginasMicrotúbulosJose AcostaAún no hay calificaciones
- BQF - 1A, B - BIOLOGIA - U IIIbDocumento31 páginasBQF - 1A, B - BIOLOGIA - U IIIbNahomy BustillosAún no hay calificaciones
- CitoesqueletoDocumento102 páginasCitoesqueletoMarco David Meza SerpaAún no hay calificaciones
- CitoesqueletoDocumento41 páginasCitoesqueletokremenchutzkyAún no hay calificaciones
- Ensayo Biología GeneralDocumento5 páginasEnsayo Biología GeneralRené VillanuevaAún no hay calificaciones
- Cito PlasmaDocumento21 páginasCito PlasmaBarbara VegaAún no hay calificaciones
- Que Son Los Microtubulos Filamentos IntermediosDocumento6 páginasQue Son Los Microtubulos Filamentos IntermediosMarvinRiveraAún no hay calificaciones
- Citoplasma y Organulos de Sintesis.Documento7 páginasCitoplasma y Organulos de Sintesis.Ninoska MartinezAún no hay calificaciones
- 6 Ensayo CitoesqueletoDocumento6 páginas6 Ensayo CitoesqueletoValeria Fernanda Rocha MerinoAún no hay calificaciones
- Citoesqueleto: FuncionesDocumento14 páginasCitoesqueleto: Funcioneskarina ramosAún no hay calificaciones
- CitoesqueletoDocumento11 páginasCitoesqueletoGabriela PeñaAún no hay calificaciones
- Citoesqueleto UPAODocumento94 páginasCitoesqueleto UPAOAndreaAún no hay calificaciones
- Tema 8 Orgánulos Celulares IDocumento16 páginasTema 8 Orgánulos Celulares Ianon_970494402Aún no hay calificaciones
- Soporte, Locomocion, Almacenamiento y Reciclado CelularDocumento36 páginasSoporte, Locomocion, Almacenamiento y Reciclado CelularLibny Quispe SegoviaAún no hay calificaciones
- Tarea 14Documento8 páginasTarea 14Paula LopezAún no hay calificaciones
- El CitoesqueletoDocumento59 páginasEl CitoesqueletoSantiago BOSOKA RIBUSAAún no hay calificaciones
- Citoesqueleto - Biología 3Documento8 páginasCitoesqueleto - Biología 3Esteban Barrios RodriguezAún no hay calificaciones
- EL CITOesqueletoDocumento4 páginasEL CITOesqueletoDiana Del Pilar Chaustre DuranAún no hay calificaciones
- MaterialComplementario Sesion05Documento20 páginasMaterialComplementario Sesion05Martin Orlando Cabrera BacaAún no hay calificaciones
- Citoesqueleto 1Documento18 páginasCitoesqueleto 1Santiago PalaciosAún no hay calificaciones
- Citosol, Comunicaciones y CitoesqueletoDocumento4 páginasCitosol, Comunicaciones y CitoesqueletoPaulaAún no hay calificaciones
- El CitoesqueletoDocumento7 páginasEl CitoesqueletoKateryn ÁlvarezAún no hay calificaciones
- Resumen Del Capitulo 15 XD SiganDocumento3 páginasResumen Del Capitulo 15 XD SiganFernanda BarzolaAún no hay calificaciones
- Taller 1. Biologia Celular y MolecularDocumento2 páginasTaller 1. Biologia Celular y MolecularMARIA LUCIA PIESCHACÓN CHUZCANOAún no hay calificaciones
- Cito EsqueletoDocumento4 páginasCito EsqueletoMateo DuraznoAún no hay calificaciones
- Citoesqueleto: Estructura y funciones clave de los microfilamentos, filamentos intermedios y microtúbulosDocumento5 páginasCitoesqueleto: Estructura y funciones clave de los microfilamentos, filamentos intermedios y microtúbulosErika Maribel BadilloAún no hay calificaciones
- Citoesqueleto PDocumento6 páginasCitoesqueleto P1900481019Aún no hay calificaciones
- 8 - Trabajo de Biologia (Word) - El CitoesqueletoDocumento3 páginas8 - Trabajo de Biologia (Word) - El CitoesqueletoAndrea Blanco Sánchez100% (1)
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
- Unidad I. Los Siete Principios de La EcologiaDocumento4 páginasUnidad I. Los Siete Principios de La EcologiaDaiana tabordaAún no hay calificaciones
- El Aparato DigestivoDocumento46 páginasEl Aparato DigestivoDaiana tabordaAún no hay calificaciones
- 1clase Introduccion 1Documento45 páginas1clase Introduccion 1Lisave CorredorAún no hay calificaciones
- Digestivo Vane-AnaDocumento18 páginasDigestivo Vane-AnaDaiana tabordaAún no hay calificaciones
- Vegetal SueloDocumento5 páginasVegetal SueloDaiana tabordaAún no hay calificaciones
- A DIFERENCIACIONES de La MEMBRANA Pagina 2019Documento54 páginasA DIFERENCIACIONES de La MEMBRANA Pagina 2019Daiana tabordaAún no hay calificaciones
- Flor e Inflorecencia 6.1Documento19 páginasFlor e Inflorecencia 6.1Daiana tabordaAún no hay calificaciones
- Anatomia y Fisiologia CirculatoriaDocumento106 páginasAnatomia y Fisiologia CirculatoriaDANIEL ALEJANDROAún no hay calificaciones
- Sistema DigestivoDocumento18 páginasSistema DigestivoDaiana tabordaAún no hay calificaciones
- Aparato CardiovascularDocumento37 páginasAparato CardiovascularAdillene Hernandez RodriguezAún no hay calificaciones
- VEGETAL3Documento2 páginasVEGETAL3Daiana tabordaAún no hay calificaciones
- Clase de Animal - Esponja-Ausencia de Columa VertebralDocumento2 páginasClase de Animal - Esponja-Ausencia de Columa VertebralDaiana tabordaAún no hay calificaciones
- Unidad I. Los Siete Principios de La EcologiaDocumento4 páginasUnidad I. Los Siete Principios de La EcologiaDaiana tabordaAún no hay calificaciones
- Tema 13b - Membranas Transporte Farmacia PDFDocumento17 páginasTema 13b - Membranas Transporte Farmacia PDFmerliaspirosgAún no hay calificaciones
- 02 Composicion Quimica de Los Seres Vivos IDocumento35 páginas02 Composicion Quimica de Los Seres Vivos IDaiana tabordaAún no hay calificaciones
- Clase de Animal - Esponja-Ausencia de Columa VertebralDocumento2 páginasClase de Animal - Esponja-Ausencia de Columa VertebralDaiana tabordaAún no hay calificaciones
- 132691059Documento18 páginas132691059Walther Smith Chasquero HuamánAún no hay calificaciones
- Clase de Celenterados: Características y TiposDocumento3 páginasClase de Celenterados: Características y TiposDaiana tabordaAún no hay calificaciones
- Fecundacion-1Documento16 páginasFecundacion-1Daiana tabordaAún no hay calificaciones
- 2 Introduccion PDFDocumento61 páginas2 Introduccion PDFYens W. Solórzano SánchezAún no hay calificaciones
- Clase de Quimica 13 de MayoDocumento2 páginasClase de Quimica 13 de MayoDaiana tabordaAún no hay calificaciones
- 10 - 8 Morfologia Tallo y AdaptacionesDocumento10 páginas10 - 8 Morfologia Tallo y Adaptacionesvodoochild_Ar80Aún no hay calificaciones
- Bacterias Vibrio y PseudomonasDocumento4 páginasBacterias Vibrio y PseudomonasDaiana tabordaAún no hay calificaciones
- T14 C Urea PDFDocumento16 páginasT14 C Urea PDFSheila EsRaAún no hay calificaciones
- Cuaderno 1 PDFDocumento68 páginasCuaderno 1 PDFIsabel AmpueroAún no hay calificaciones
- 9264-Texto Del Artículo-28222-1-10-20190412 PDFDocumento12 páginas9264-Texto Del Artículo-28222-1-10-20190412 PDFJohnva_92Aún no hay calificaciones
- 2 Introduccion PDFDocumento61 páginas2 Introduccion PDFYens W. Solórzano SánchezAún no hay calificaciones
- Propuesta para Un Vocabulario Mínimo en CienciaDocumento8 páginasPropuesta para Un Vocabulario Mínimo en CienciaDaiana tabordaAún no hay calificaciones
- Lamarck DarwinDocumento20 páginasLamarck DarwinGuille Pineda100% (1)
- Clasificación de zonas de vida en el PerúDocumento30 páginasClasificación de zonas de vida en el PerúAnonymous iGoIeWrH6J100% (1)
- Gestión residuos sólidosDocumento18 páginasGestión residuos sólidosKatherine Jurado ToledoAún no hay calificaciones
- ECOGRAFIADocumento12 páginasECOGRAFIAElianAún no hay calificaciones
- Modelo PosterDocumento1 páginaModelo PosterRoyer YampierAún no hay calificaciones
- Tema - 12. Audesirk 2013-651-671 Circulac PDFDocumento21 páginasTema - 12. Audesirk 2013-651-671 Circulac PDFMarco Joel Pinto MaidanaAún no hay calificaciones
- Manual de Agentes FísicosDocumento17 páginasManual de Agentes FísicosMariana CaballeroAún no hay calificaciones
- Quemaduras TérmicasDocumento37 páginasQuemaduras TérmicasMariel CuelloAún no hay calificaciones
- Efecto pirita lixiviación calcopiritaDocumento12 páginasEfecto pirita lixiviación calcopiritaJose Antonio RosasAún no hay calificaciones
- Tema 1 BiologiaDocumento28 páginasTema 1 BiologiaMoonlight Notes moonlightstudygramAún no hay calificaciones
- Evaluación de Física - Conversión - Notación CientíficaDocumento2 páginasEvaluación de Física - Conversión - Notación CientíficaROMINA ADAROAún no hay calificaciones
- CuecueyosDocumento16 páginasCuecueyosivan cantoAún no hay calificaciones
- Dieta AbcdeDocumento15 páginasDieta AbcdeJuan PawneesAún no hay calificaciones
- AcetilcolinaDocumento39 páginasAcetilcolinamickevillanuevaAún no hay calificaciones
- Semana 06 SimulacroDocumento8 páginasSemana 06 SimulacroTotti Di Franchesco PelaezAún no hay calificaciones
- Salida A SogayDocumento3 páginasSalida A Sogaysofia umpireAún no hay calificaciones
- Manual de Recomendaciones Sobre Sistemas Intensivos de Producción de Carne-MinDocumento58 páginasManual de Recomendaciones Sobre Sistemas Intensivos de Producción de Carne-MinJose Rodrigo Rojas GonzalezAún no hay calificaciones
- Practica No5.Celomadso-EnterocelomadosDocumento16 páginasPractica No5.Celomadso-EnterocelomadosAlan DavidAún no hay calificaciones
- Libro Ciclo Repaso 2014Documento213 páginasLibro Ciclo Repaso 2014Santiago Sánchez100% (1)
- Benford, Gregory - Saga Del Centro GalacticoDocumento3 páginasBenford, Gregory - Saga Del Centro GalacticoJorge Andres NicolaldeAún no hay calificaciones
- Citogenética Expo CompletoDocumento56 páginasCitogenética Expo CompletoPrincesita Kerli DelgadoAún no hay calificaciones
- Ecologia y Ecosistemas Semana 2Documento30 páginasEcologia y Ecosistemas Semana 2Jhonny AlexanderAún no hay calificaciones
- Unidad 1 Administracion EstrategicaDocumento5 páginasUnidad 1 Administracion Estrategicakerin_lmsAún no hay calificaciones
- HolaDocumento13 páginasHolaEspresso NotdepressoAún no hay calificaciones
- AFC Guia Planificacion Predial Pagina BajaDocumento56 páginasAFC Guia Planificacion Predial Pagina BajaWilliam Passo Peinado100% (1)
- El Sistema de Palancas Base para El Analisis Mecanico Del Movimiento Corporal Humano Y Sus AlteracionesDocumento12 páginasEl Sistema de Palancas Base para El Analisis Mecanico Del Movimiento Corporal Humano Y Sus AlteracionesEva CarbajalAún no hay calificaciones
- Leyes de Mendel y sus experimentos con guisantesDocumento5 páginasLeyes de Mendel y sus experimentos con guisantesMaria Victoria SpinettoAún no hay calificaciones
- La Rebelión de Los Brujos - Louis Pauwells & Jacques BergierDocumento809 páginasLa Rebelión de Los Brujos - Louis Pauwells & Jacques BergierCarme CameAún no hay calificaciones
- Planificación Anual de Ciencias Naturales 2022Documento10 páginasPlanificación Anual de Ciencias Naturales 2022Karina Julia Tirasso100% (2)
- Atlas Artropodos Vulnerables Tcm30-198227Documento721 páginasAtlas Artropodos Vulnerables Tcm30-198227Nacho GallentAún no hay calificaciones