También podría gustarte
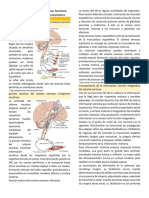
- Cap 45 Organización Del Sistema Nervioso, Funciones Básicas de Las Sinapsis y NeurotransmisoresDocumento11 páginasCap 45 Organización Del Sistema Nervioso, Funciones Básicas de Las Sinapsis y Neurotransmisoresrodrigo4590100% (2)
- Apuntes Sinapsis 1Documento10 páginasApuntes Sinapsis 1Vanessa MuñozAún no hay calificaciones
- CLASE 1 y 2Documento19 páginasCLASE 1 y 2lilianaAún no hay calificaciones
- Cap. 46 Organizacion Del Sistema NerviosoDocumento12 páginasCap. 46 Organizacion Del Sistema NerviosoYulia Gomez100% (1)
- Capítulo 46Documento4 páginasCapítulo 46Cynthia GonzálezAún no hay calificaciones
- 01 Diseño General Del SNCDocumento18 páginas01 Diseño General Del SNCElvis SantayAún no hay calificaciones
- Organización Del Sistema Nervioso, Funciones Básicas de La Sinapsis y NeurotransmisoresDocumento21 páginasOrganización Del Sistema Nervioso, Funciones Básicas de La Sinapsis y NeurotransmisoresGonzalo Córdova OrtegaAún no hay calificaciones
- Resumen Capitulo 46Documento4 páginasResumen Capitulo 46Rashel MoscosoAún no hay calificaciones
- Informe NeuroanatomiaDocumento18 páginasInforme NeuroanatomiaMarjorie Dayana JaponAún no hay calificaciones
- Cap46 Organizacion Del Sistema NerviosoDocumento12 páginasCap46 Organizacion Del Sistema NerviosodavidmoralesAún no hay calificaciones
- Sensibilidades Somaticas I Organizacion General de Las Sensaciones Tactil y PosicionalDocumento41 páginasSensibilidades Somaticas I Organizacion General de Las Sensaciones Tactil y PosicionalRuddy Morales FloresAún no hay calificaciones
- Unidad IX: El Sistema Nervioso: A. Principios Generales y Fisiología de La SensibilidadDocumento8 páginasUnidad IX: El Sistema Nervioso: A. Principios Generales y Fisiología de La SensibilidadMaribel Alvarez de la EspriellaAún no hay calificaciones
- Anatomia y Fisiologia Sistema NerviosoDocumento10 páginasAnatomia y Fisiologia Sistema NerviosoCamila Alvarez MayorgaAún no hay calificaciones
- Niveles NerviososDocumento5 páginasNiveles NerviososangyAún no hay calificaciones
- Sistema NerviosoDocumento12 páginasSistema NerviosoFaisy AlcántaraAún no hay calificaciones
- DocumentoDocumento4 páginasDocumentoCatalina CacaceAún no hay calificaciones
- Sandoval Jennifher UTHDocumento11 páginasSandoval Jennifher UTHEstrella MaldonadoAún no hay calificaciones
- Fisiologia SNCDocumento18 páginasFisiologia SNCAndrea Narvaez UribeAún no hay calificaciones
- Clase 2 Anatomia y Fisiologia Del SNDocumento2 páginasClase 2 Anatomia y Fisiologia Del SNBarby RodriguezAún no hay calificaciones
- Tema 2 - Sistema Nervioso - Fleming - Version Definitiva PDFDocumento27 páginasTema 2 - Sistema Nervioso - Fleming - Version Definitiva PDFCARLOS COSTASAún no hay calificaciones
- DISEnO GENERAL DEL SISTEMA NERVIOSODocumento26 páginasDISEnO GENERAL DEL SISTEMA NERVIOSOwilly joelAún no hay calificaciones
- Taller 1 Sistema NerviosoDocumento6 páginasTaller 1 Sistema NerviosoInesita Ortega Silva100% (2)
- Tarea 3 AnatomiaDocumento5 páginasTarea 3 AnatomiaEridania Delgado DiazAún no hay calificaciones
- Bases Biologicas Del Comportamiento Humano Psicologia General Parte IIIDocumento4 páginasBases Biologicas Del Comportamiento Humano Psicologia General Parte IIIjesusAún no hay calificaciones
- Farmaco SNCDocumento7 páginasFarmaco SNCYass MuñozAún no hay calificaciones
- Sistema Nervioso PerifericoDocumento14 páginasSistema Nervioso PerifericoCarlos Arana MendezAún no hay calificaciones
- Sistema Nervioso PeriféricoDocumento2 páginasSistema Nervioso PeriféricoDiego Francisco Mendoza FriasAún no hay calificaciones
- Capítulo 46. Organización Del Sistema Nervioso, Funciones Básicas de Las Sinapsis y NeurotransmisoresDocumento7 páginasCapítulo 46. Organización Del Sistema Nervioso, Funciones Básicas de Las Sinapsis y NeurotransmisoresJhony PradelaAún no hay calificaciones
- Resumen Fisiología - Organización Del Sistema NerviosoDocumento17 páginasResumen Fisiología - Organización Del Sistema NerviosoAntonella ArdilesAún no hay calificaciones
- Endocrino y NerviosoDocumento24 páginasEndocrino y NerviosoYesenia Andrea Martinez HerreraAún no hay calificaciones
- Taller 1 Sistema NerviosoDocumento4 páginasTaller 1 Sistema NerviosoNeon LogAún no hay calificaciones
- Sistema Nervioso y PeriféricoDocumento10 páginasSistema Nervioso y PeriféricocristhinaAún no hay calificaciones
- Sistema Nervioso Central Y PeriféricoDocumento3 páginasSistema Nervioso Central Y PeriféricoLuciana NapolitanoAún no hay calificaciones
- Sistema Nervioso IDocumento14 páginasSistema Nervioso IMonica MarianiAún no hay calificaciones
- Tarea 2Documento4 páginasTarea 2carlos manuel figueroaAún no hay calificaciones
- FisiologíaDocumento15 páginasFisiologíaMaura MeloAún no hay calificaciones
- Taller de Biologia Grado 8Documento4 páginasTaller de Biologia Grado 8Oscar Fernando Portilla GómezAún no hay calificaciones
- Material Del Sistema NerviosoDocumento6 páginasMaterial Del Sistema Nerviosoalmenar24Aún no hay calificaciones
- Farmacología y Toxicología SNDocumento60 páginasFarmacología y Toxicología SNNelson Mandela Ramos AlonsoAún no hay calificaciones
- .E.A. Guido VillagomezDocumento5 páginas.E.A. Guido VillagomezMarcelo ChavezAún no hay calificaciones
- SNC - Grupo 6 - Fisiologia IiDocumento43 páginasSNC - Grupo 6 - Fisiologia Iirutmeraz01Aún no hay calificaciones
- NeurotoxicologíaDocumento44 páginasNeurotoxicologíaGrety Huaycama SangamaAún no hay calificaciones
- Clase Capitulo 3 Anatomia Del Sistema NerviosoDocumento43 páginasClase Capitulo 3 Anatomia Del Sistema NerviosoAnónimo9Aún no hay calificaciones
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Bases Biologicas Del ComportamientoDocumento36 páginasBases Biologicas Del Comportamientolaura100% (2)
- Bases Neuroficiologicas Del AprendizajeDocumento6 páginasBases Neuroficiologicas Del AprendizajeNatalia FariñaAún no hay calificaciones
- Capítulo 45 Organización Del Sistema Nervioso Funciones Sinapsis y NeurotransmisoresDocumento7 páginasCapítulo 45 Organización Del Sistema Nervioso Funciones Sinapsis y NeurotransmisoresDiego Rios CaballeroAún no hay calificaciones
- Sistema NerviosoDocumento32 páginasSistema NerviosoqfbedithcdzAún no hay calificaciones
- T. 2 Fundamentos BiológicosDocumento9 páginasT. 2 Fundamentos BiológicosJulia QuismondoAún no hay calificaciones
- Sistema NerviosoDocumento44 páginasSistema NerviosoDebo GarciaAún no hay calificaciones
- Resumen 2do ParcialDocumento18 páginasResumen 2do ParcialmalAún no hay calificaciones
- Taller No 6Documento5 páginasTaller No 6api-241769574Aún no hay calificaciones
- Resumen Capitulo 46 GuytonDocumento8 páginasResumen Capitulo 46 GuytonMcPotatoe :86% (7)
- Biologia Guia de EstudioDocumento9 páginasBiologia Guia de EstudiohscarvajalsAún no hay calificaciones
- Taller 1 Sistema NerviosoDocumento5 páginasTaller 1 Sistema NerviosoRobertoCarlosHmaJimenezZappaAún no hay calificaciones
- Anatomía SNC bioTKDocumento11 páginasAnatomía SNC bioTKMarii Alejandra MartiinezAún no hay calificaciones
- Cuadro Comparativo PresentarDocumento4 páginasCuadro Comparativo PresentarLuz Mollo F.Aún no hay calificaciones
- Sistema NerviosoDocumento26 páginasSistema NerviosodatanaAún no hay calificaciones
- Trabajo Final AnatomiaDocumento14 páginasTrabajo Final AnatomiaAna Marte GAún no hay calificaciones
- Conferencia 5Documento28 páginasConferencia 5Daniel AjilaAún no hay calificaciones
- Desarrolo HumanoDocumento11 páginasDesarrolo HumanoOmar VegaAún no hay calificaciones
- Memoria PTTDocumento21 páginasMemoria PTTOmar VegaAún no hay calificaciones
- Desarrolo HumanoDocumento11 páginasDesarrolo HumanoOmar VegaAún no hay calificaciones
- Indefension AprendidaDocumento7 páginasIndefension AprendidaOmar VegaAún no hay calificaciones
- PDFDocumento11 páginasPDFben_esmAún no hay calificaciones
- Ensayo MemoriaDocumento7 páginasEnsayo MemoriaOmar VegaAún no hay calificaciones
- Distorciones y Alteracions de La MemoriaDocumento5 páginasDistorciones y Alteracions de La MemoriaOmar VegaAún no hay calificaciones
- Mecanismos de Defensa PresentacionDocumento8 páginasMecanismos de Defensa PresentacionOmar VegaAún no hay calificaciones
- Organizacion Del Sistema Nervioso PPDocumento8 páginasOrganizacion Del Sistema Nervioso PPOmar VegaAún no hay calificaciones
- La Maldicion de Conocimiento PTTDocumento5 páginasLa Maldicion de Conocimiento PTTOmar VegaAún no hay calificaciones
- Trastorno BipolarDocumento16 páginasTrastorno BipolarOmar VegaAún no hay calificaciones
- Indefension AprendidaDocumento7 páginasIndefension AprendidaOmar VegaAún no hay calificaciones
- Terapias PsicologicasDocumento4 páginasTerapias PsicologicasOmar VegaAún no hay calificaciones
- Dunning KrugerDocumento4 páginasDunning KrugerOmar VegaAún no hay calificaciones
- La Maldicion Del ConocimientoDocumento5 páginasLa Maldicion Del ConocimientoOmar VegaAún no hay calificaciones
- El Proceso de Convertise en PersonaDocumento4 páginasEl Proceso de Convertise en PersonaOmar VegaAún no hay calificaciones
- Etapas Del Desarrollo Del Sistema NerviosoDocumento11 páginasEtapas Del Desarrollo Del Sistema NerviosoOmar VegaAún no hay calificaciones
- Distorciones y Alteracions de La MemoriaDocumento5 páginasDistorciones y Alteracions de La MemoriaOmar VegaAún no hay calificaciones
- Ensayo MemoriaDocumento7 páginasEnsayo MemoriaOmar VegaAún no hay calificaciones
- Informe de Sintesis de La EspinelaDocumento8 páginasInforme de Sintesis de La EspinelaMarlen FuentesAún no hay calificaciones
- Fundamentos de Las Operaciones Unitarias de Transferencia de MasaDocumento21 páginasFundamentos de Las Operaciones Unitarias de Transferencia de MasaJorge VasquezAún no hay calificaciones
- DibenzalacetonaDocumento10 páginasDibenzalacetonaMaria100% (1)
- Mishel Naranjo BF03 Tarea 1Documento2 páginasMishel Naranjo BF03 Tarea 1Mishel NaranjoAún no hay calificaciones
- Programa Analítico de QuímicaDocumento2 páginasPrograma Analítico de QuímicaRITA BARRIONUEVOAún no hay calificaciones
- Fisicoquimica InfografiaDocumento1 páginaFisicoquimica InfografiaMonse CoreyAún no hay calificaciones
- POE de ALMACENAMIENTO PARA bOTICADocumento6 páginasPOE de ALMACENAMIENTO PARA bOTICAbeto100% (2)
- Bioquimica Oral Placa MicrobianaDocumento14 páginasBioquimica Oral Placa MicrobianaCristian Pincheira ArayaAún no hay calificaciones
- Laboratorio de Quimica (Medicion de Volumen)Documento7 páginasLaboratorio de Quimica (Medicion de Volumen)Evelyn DiazAún no hay calificaciones
- Salicilato de MetiloDocumento3 páginasSalicilato de MetiloMadeleine RodriguezAún no hay calificaciones
- Informe Instalación de TransductoresDocumento20 páginasInforme Instalación de TransductoresHarold EnriqueAún no hay calificaciones
- Investigacion Documental .Metodos DirectosDocumento11 páginasInvestigacion Documental .Metodos DirectosYARETZI GUADALUPE MENDOZA GARCIAAún no hay calificaciones
- Formato Cuadro Comparativo Actividad 5Documento8 páginasFormato Cuadro Comparativo Actividad 5KlderonLemus0% (1)
- All Be Gone TDS CT-217 (Spanish)Documento1 páginaAll Be Gone TDS CT-217 (Spanish)Sergio TrevinoAún no hay calificaciones
- Ejercicios Sobre SolucionesDocumento6 páginasEjercicios Sobre SolucionescristinaAún no hay calificaciones
- Curva de Valoración de Yodo Con Tiosulfato de SodioDocumento3 páginasCurva de Valoración de Yodo Con Tiosulfato de SodioRodríguez Vázquez EvelynAún no hay calificaciones
- Resume NDocumento6 páginasResume NDeyvis ChihuanAún no hay calificaciones
- El PerfumeDocumento11 páginasEl Perfumegbarrio qAún no hay calificaciones
- Informe Concepto Del MolDocumento6 páginasInforme Concepto Del MolSteven CastilloAún no hay calificaciones
- Lixiviación en BateasDocumento10 páginasLixiviación en BateasJoaquin Perez GonzalezAún no hay calificaciones
- Reducción de Ésteres y Ácidos CarboxílicosDocumento5 páginasReducción de Ésteres y Ácidos CarboxílicosAndrés VelázquezAún no hay calificaciones
- Trabajo de QuimicaDocumento10 páginasTrabajo de QuimicaKlk Maoly JsjsjAún no hay calificaciones
- 6 Plan Mejoramiento 2SDocumento3 páginas6 Plan Mejoramiento 2Semiliano04cubillosAún no hay calificaciones
- Practica N 4 Equilibrio Quimico PrincipiDocumento9 páginasPractica N 4 Equilibrio Quimico Principicatalina.olivosAún no hay calificaciones
- HDS TippsLine Limpia Vidrios V1Documento8 páginasHDS TippsLine Limpia Vidrios V1tatianaAún no hay calificaciones
- Funciones de ProteinasDocumento35 páginasFunciones de ProteinasDx LuffyAún no hay calificaciones
- El Sistema Respiratorio para Segundo de PrimariaDocumento4 páginasEl Sistema Respiratorio para Segundo de PrimariaRO PEREZ TREJOAún no hay calificaciones
- T4 Plasticos Impresion3dDocumento7 páginasT4 Plasticos Impresion3dCharlie A. BadilloAún no hay calificaciones
- Química Orgánica Clase 1. GeneralidadesDocumento54 páginasQuímica Orgánica Clase 1. GeneralidadesFelipe A. PérezAún no hay calificaciones
- Mezcla de GasesDocumento15 páginasMezcla de GasesCris HerreraAún no hay calificaciones