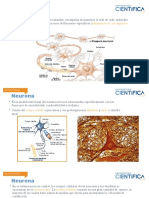
También podría gustarte
- wuolah-free-TEMA-7-FISIOLOGIA NNNNNNNDocumento6 páginaswuolah-free-TEMA-7-FISIOLOGIA NNNNNNNRodrigo Vallecillo GrauAún no hay calificaciones
- Notas NeuropsicologíaDocumento20 páginasNotas NeuropsicologíaSara OchoaAún no hay calificaciones
- Función Del Sistema Nervioso MotorDocumento1 páginaFunción Del Sistema Nervioso Motor20-v.lopez.oAún no hay calificaciones
- Mapa MentalDocumento5 páginasMapa MentalViviana Maribi Morocho RíosAún no hay calificaciones
- Wuolah Free Tema 2 y 3Documento14 páginasWuolah Free Tema 2 y 3Monica Quero FernandezAún no hay calificaciones
- Tema 2 NeurogastronomiaDocumento53 páginasTema 2 NeurogastronomiaAlejandra TafurAún no hay calificaciones
- Cuaderno de Practicas Bases Biologicas Semana 5 6 y 7Documento9 páginasCuaderno de Practicas Bases Biologicas Semana 5 6 y 7Emerson Panuera GAún no hay calificaciones
- Maria Morel 10138679Documento1 páginaMaria Morel 10138679Maria MorelAún no hay calificaciones
- Tarea #7Documento3 páginasTarea #7lrutyAún no hay calificaciones
- Clase 13 NEURO DB PDF AulaDocumento36 páginasClase 13 NEURO DB PDF AulaEdson ChaucaGarciaAún no hay calificaciones
- Semana 2 Enfoques de La NeuropsicologiaDocumento3 páginasSemana 2 Enfoques de La NeuropsicologiaMuriel BandaAún no hay calificaciones
- Diagrama IshikawaDocumento4 páginasDiagrama IshikawaIzabethel VargasAún no hay calificaciones
- Wuolah Free Tema5Documento24 páginasWuolah Free Tema5Angie ValenciaAún no hay calificaciones
- Biología Del Aprendizaje 1.1.Documento31 páginasBiología Del Aprendizaje 1.1.gisela magali torresAún no hay calificaciones
- Semana 13Documento19 páginasSemana 13Rony CastañedaAún no hay calificaciones
- Clase 3 Sinapsis-Y-Union-NeuromuscularDocumento25 páginasClase 3 Sinapsis-Y-Union-NeuromuscularAndres BorborAún no hay calificaciones
- Organización Del Sistema Nervioso, Funciones Básicas de Las Sinapsis y Neurotransmisores.Documento36 páginasOrganización Del Sistema Nervioso, Funciones Básicas de Las Sinapsis y Neurotransmisores.Manuel Miranda100% (5)
- Anatomia, Tarea 7Documento7 páginasAnatomia, Tarea 7Mariel FernandezAún no hay calificaciones
- Tarea 1 de AnatomiaDocumento6 páginasTarea 1 de AnatomiaAna AbadAún no hay calificaciones
- Wuolah-Free-Tema 8 AprendizajeDocumento9 páginasWuolah-Free-Tema 8 AprendizajeluciaAún no hay calificaciones
- Neurofisiología de La Respuesta Sexual Humana Cano Martinez AndreaDocumento31 páginasNeurofisiología de La Respuesta Sexual Humana Cano Martinez AndreaAndrea Cano MartinezAún no hay calificaciones
- NeurotransmisoresDocumento25 páginasNeurotransmisoresJair Alexis Chavez100% (2)
- 60 Esquemas MédicosDocumento61 páginas60 Esquemas MédicosVALERIA MARIN MARTINEZ100% (2)
- Cap 46 GuytonDocumento8 páginasCap 46 GuytonCinthia Lemus100% (1)
- BloquesDocumento3 páginasBloquesCamila PastorAún no hay calificaciones
- Tema 4. MOVIMIENTODocumento74 páginasTema 4. MOVIMIENTOlaugil888Aún no hay calificaciones
- Sesión 2 en TextoDocumento4 páginasSesión 2 en TextoINGRID ETEL LOPEZ DEL AGUILAAún no hay calificaciones
- Psicología T.2 Bases Biológicas de La Conducta HumanaDocumento10 páginasPsicología T.2 Bases Biológicas de La Conducta Humanaebarriose12221Aún no hay calificaciones
- TEMA 2 PSICOlogiaDocumento24 páginasTEMA 2 PSICOlogiaGeomate Reforzamiento y Asesoría AcadémicaAún no hay calificaciones
- Clase #1Documento50 páginasClase #1fernandafigueroa2012Aún no hay calificaciones
- Regulación Del Movimiento, Papel de Los Ganglios Basales y Cerebelo 25.01.22Documento22 páginasRegulación Del Movimiento, Papel de Los Ganglios Basales y Cerebelo 25.01.22oskitar0123Aún no hay calificaciones
- GuytonDocumento8 páginasGuytonhelenaserrano15Aún no hay calificaciones
- NeurocienciaDocumento1 páginaNeurocienciaSara RochaAún no hay calificaciones
- Free Tema 5.2Documento7 páginasFree Tema 5.2ANA SOFÍA MORALES RODRÍGUEZAún no hay calificaciones
- Los 10 Neurotransmisores Principales y Su Función en El Sistema Nervioso CentralDocumento8 páginasLos 10 Neurotransmisores Principales y Su Función en El Sistema Nervioso Centralrosanalinda20104Aún no hay calificaciones
- APUNTES MPGS. Trastornos de SueñoDocumento20 páginasAPUNTES MPGS. Trastornos de SueñoVioleta Medialdea MorillasAún no hay calificaciones
- Anatomia Tarea 1Documento3 páginasAnatomia Tarea 1Ocalys Hiciano57% (7)
- Clase 03Documento64 páginasClase 03BRIGGITTE QUISPEEAún no hay calificaciones
- Wuolah Free Modelo Control Motor15Documento30 páginasWuolah Free Modelo Control Motor15Mary GomezAún no hay calificaciones
- Neurona y Funciones Superiores LGDocumento39 páginasNeurona y Funciones Superiores LGValentina ZamudioAún no hay calificaciones
- 10 - Organización Del Sistema NerviosoDocumento5 páginas10 - Organización Del Sistema NerviosoCeleste GuanaisAún no hay calificaciones
- Capturas Imágenes de Biotk - Mercy - Timote - Grupo - 17Documento4 páginasCapturas Imágenes de Biotk - Mercy - Timote - Grupo - 17Julian Alberto Gonzalez TorresAún no hay calificaciones
- Cerebro, Neurona, Apego y DrogasDocumento8 páginasCerebro, Neurona, Apego y DrogasAbigailAún no hay calificaciones
- Sistema NerviosoDocumento26 páginasSistema NerviosovenusciteresAún no hay calificaciones
- Sistema Nervioso Parte IDocumento16 páginasSistema Nervioso Parte IIgnacio NavarroAún no hay calificaciones
- Repaso NeurofisioDocumento7 páginasRepaso NeurofisioESTER ABIGAIL ZECEÑA ZAPETAún no hay calificaciones
- UD1 El Control MotorDocumento52 páginasUD1 El Control MotorDavid Obeso VicenteAún no hay calificaciones
- Estados de Actividad Cerebral Sistema Nervioso VegetativoDocumento65 páginasEstados de Actividad Cerebral Sistema Nervioso VegetativoXIOMARA TOPACIO LEVANO BELLIDOAún no hay calificaciones
- ACTIVIDAD RETADORA No2 DDDDDDocumento2 páginasACTIVIDAD RETADORA No2 DDDDDAnaly Mamani TorresAún no hay calificaciones
- TAREA 7. Anatomia y FisiologiaDocumento6 páginasTAREA 7. Anatomia y FisiologiaCamila CruzAún no hay calificaciones
- EVA 1 PSICOMOTRICIDAD TerminadoDocumento5 páginasEVA 1 PSICOMOTRICIDAD TerminadoLucero Auriestela Rivera HernandezAún no hay calificaciones
- Clase 4 - Neurologia UnisonDocumento91 páginasClase 4 - Neurologia Unisonsol sanchezAún no hay calificaciones
- Introducción Al Sistema Nervioso ZootecniaDocumento34 páginasIntroducción Al Sistema Nervioso ZootecniaSEBASTIAN TAMAYO LONDONOAún no hay calificaciones
- Principios Básicos de Neurofisiología 1Documento18 páginasPrincipios Básicos de Neurofisiología 1Erika Moreno de León100% (1)
- Neurociencias en El Contexto MentalDocumento20 páginasNeurociencias en El Contexto MentalYuliana GarzonAún no hay calificaciones
- Funciones Corticales 2015Documento6 páginasFunciones Corticales 2015Matias JulianAún no hay calificaciones
- Diagnóstico de Alteraciones Temporomandibulares Semana 4Documento2 páginasDiagnóstico de Alteraciones Temporomandibulares Semana 4cd.brendasotofloresAún no hay calificaciones
- Depresión y NeuroplasticidadDocumento115 páginasDepresión y NeuroplasticidadAntero Vasquez MejiaAún no hay calificaciones
- Mapa 1Documento1 páginaMapa 1camilaAún no hay calificaciones
- wuolah-free-TEMA 6Documento15 páginaswuolah-free-TEMA 6Angie ValenciaAún no hay calificaciones
- Wuolah Free Tema5Documento24 páginasWuolah Free Tema5Angie ValenciaAún no hay calificaciones
- wuolah-free-TEMA 7PSDocumento9 páginaswuolah-free-TEMA 7PSAngie ValenciaAún no hay calificaciones
- Pensamiento. Como Ya Se Ha Mencionado en ElDocumento12 páginasPensamiento. Como Ya Se Ha Mencionado en ElAngie ValenciaAún no hay calificaciones
- UntitledDocumento24 páginasUntitledAngie ValenciaAún no hay calificaciones
- Luchando Contra El COVID-19 Desinformación Sobre Redes Sociales: Evidencia Experimental de Una Intervención Escalable de Precisión y EmpujónDocumento11 páginasLuchando Contra El COVID-19 Desinformación Sobre Redes Sociales: Evidencia Experimental de Una Intervención Escalable de Precisión y EmpujónAngie ValenciaAún no hay calificaciones
- Banco de Preguntas Enfermedades NeurodegenerativasDocumento14 páginasBanco de Preguntas Enfermedades Neurodegenerativasdamaris jocelyn cipres vallejoAún no hay calificaciones
- Primer Examen Parcial de Biología Celular UPIBIDocumento16 páginasPrimer Examen Parcial de Biología Celular UPIBIKelly Ortiz FrancoAún no hay calificaciones
- Guía de Estudio Solemne 2Documento6 páginasGuía de Estudio Solemne 2Matías Alexis Gutiérrez PradenasAún no hay calificaciones
- Cromosoma yDocumento79 páginasCromosoma yEsther Elizabeth GualanAún no hay calificaciones
- Practicas de Biotecnologia - 2 FaseDocumento51 páginasPracticas de Biotecnologia - 2 FaseJHESSICA ETHEL PIMENTEL GONZALESAún no hay calificaciones
- Biología 4º EsoDocumento81 páginasBiología 4º EsoAlmudena Ciordia Gonzalez-AllerAún no hay calificaciones
- Aspectos Generales Del Metabolismo IntermediarioDocumento3 páginasAspectos Generales Del Metabolismo IntermediarioDaniela RoseroAún no hay calificaciones
- Neurotransmisores y Receptores-Contraccion MuscularDocumento12 páginasNeurotransmisores y Receptores-Contraccion MuscularJosé Andrés Patricio Muñoz AlbornozAún no hay calificaciones
- Taller Tabla PeriodicaDocumento2 páginasTaller Tabla PeriodicaJavier Mercado DiazAún no hay calificaciones
- 2° Medio Biología 19-03 - CelulaDocumento18 páginas2° Medio Biología 19-03 - Celulaprofesora María Antonieta TorresAún no hay calificaciones
- Genética ForenseDocumento5 páginasGenética ForenseSara Maria Ortega salasAún no hay calificaciones
- Acidos-Nucleicos PPT VeterinariaDocumento18 páginasAcidos-Nucleicos PPT VeterinariaRoller VizcarraAún no hay calificaciones
- DSCDocumento7 páginasDSCCeci SotoAún no hay calificaciones
- Manual 2020 Lab 3 RevisadoDocumento8 páginasManual 2020 Lab 3 RevisadoDiana MuñozAún no hay calificaciones
- Apuntes I - Nutricion Humana - 2023Documento22 páginasApuntes I - Nutricion Humana - 2023Jesica VillalbaAún no hay calificaciones
- HIGADO GRASO Convertido FusionadoDocumento47 páginasHIGADO GRASO Convertido FusionadoLex GerardoAún no hay calificaciones
- Etapas Del Anabolismo y CatabolismoDocumento8 páginasEtapas Del Anabolismo y CatabolismoAxel MelchorAún no hay calificaciones
- Factores Que Afectan La Actividad EnzimáticaDocumento4 páginasFactores Que Afectan La Actividad EnzimáticaDavid DiazAún no hay calificaciones
- Temas Examen Serum Julio2017Documento4 páginasTemas Examen Serum Julio2017Mitsuki LiAún no hay calificaciones
- Biologia CelularDocumento13 páginasBiologia CelularEdward ArredondoAún no hay calificaciones
- S2 PPT LaboratorioDocumento26 páginasS2 PPT LaboratorioValeria CarrionAún no hay calificaciones
- Informe de BiomoleculasDocumento6 páginasInforme de BiomoleculasAlex Roa BernalAún no hay calificaciones
- Genética - Seminario 10Documento18 páginasGenética - Seminario 10Alessandro ValdiviesoAún no hay calificaciones
- Bioquimica Practica #9 Trigliceridos IiDocumento12 páginasBioquimica Practica #9 Trigliceridos Iifrank ganz bravoAún no hay calificaciones
- Informe - BOMBEO DE PROTONES EN LEVADURAS PDFDocumento4 páginasInforme - BOMBEO DE PROTONES EN LEVADURAS PDFGustavo Gutiérrez Gómez67% (3)
- Ensayo - VitaminasDocumento7 páginasEnsayo - VitaminasEDWIN CAMBOAún no hay calificaciones
- TG Informe BCT IIDocumento8 páginasTG Informe BCT IIJouRojoLópezAún no hay calificaciones
- Reacciones Químicas de Los CarbohidratosDocumento7 páginasReacciones Químicas de Los CarbohidratosWil BardalesAún no hay calificaciones
- Proteinas Globulares y FibrosasDocumento15 páginasProteinas Globulares y Fibrosassara henao100% (1)
- GlucosidasaDocumento126 páginasGlucosidasaMaritsa PerHerAún no hay calificaciones