También podría gustarte
- Programas de Protección Regional de Áreas Prioritarias de grandes ballenas en el golfo de California y costa occidental de Baja California Sur: Propuestas de conservaciónDe EverandProgramas de Protección Regional de Áreas Prioritarias de grandes ballenas en el golfo de California y costa occidental de Baja California Sur: Propuestas de conservaciónAún no hay calificaciones
- Abundancia Estival de Aves Costeras y Mamíferos Marinos en Islas Desertores, Comuna de Chaitén (Patagonia ChilenaDocumento12 páginasAbundancia Estival de Aves Costeras y Mamíferos Marinos en Islas Desertores, Comuna de Chaitén (Patagonia Chilenaclaudio delgadoAún no hay calificaciones
- Asesinato en La Plata - 230607 - 114552Documento44 páginasAsesinato en La Plata - 230607 - 114552Vitotigre HolaAún no hay calificaciones
- Lavinia Flores-Cascante, Benjamín Morales-Vela, Nataly Castelblanco-Martínez, Janneth Padilla-Saldívar y Nicole AuilDocumento12 páginasLavinia Flores-Cascante, Benjamín Morales-Vela, Nataly Castelblanco-Martínez, Janneth Padilla-Saldívar y Nicole AuilLuisAún no hay calificaciones
- 2016 Ficha Tortuga CaguamaDocumento4 páginas2016 Ficha Tortuga CaguamaHeidi GutierrezAún no hay calificaciones
- Crustaceos FinalDocumento19 páginasCrustaceos FinalJuan FelipeAún no hay calificaciones
- Que LeleDocumento8 páginasQue LeleLucy AlmarazAún no hay calificaciones
- FT 8Documento19 páginasFT 8Ronald Rimari BarzolaAún no hay calificaciones
- Cupul-Magaña Etal 2000ComunidadescoralinasMarietasyBadeBaDocumento8 páginasCupul-Magaña Etal 2000ComunidadescoralinasMarietasyBadeBaRosa Carmen Sotelo CasasAún no hay calificaciones
- Lora Vascular y Formaciones Vegetacionales en Elsitio de Estudios Socio Ecológicos A Largo Plazo, Isla Gonzalo, Archipiélago Diego RamírezDocumento10 páginasLora Vascular y Formaciones Vegetacionales en Elsitio de Estudios Socio Ecológicos A Largo Plazo, Isla Gonzalo, Archipiélago Diego RamírezRobinAún no hay calificaciones
- Patrones de Distribución de Haemulidae A Lo Largo Del Noreste de BrasilDocumento24 páginasPatrones de Distribución de Haemulidae A Lo Largo Del Noreste de Brasilluis medina “el medis”Aún no hay calificaciones
- Herpetofauna de La Reserva Natural Provincial Isla Apipé Grande (Corrientes, Argentina)Documento8 páginasHerpetofauna de La Reserva Natural Provincial Isla Apipé Grande (Corrientes, Argentina)Karen AlmeidaAún no hay calificaciones
- Equinodermos Del Parque Nacional Isla Isabel, Nayarit, MéxicoDocumento11 páginasEquinodermos Del Parque Nacional Isla Isabel, Nayarit, MéxicoCRISTIAN MOISES GALVAN VILLAAún no hay calificaciones
- 1264-Texto Del Artículo-2061-1-10-20200304Documento15 páginas1264-Texto Del Artículo-2061-1-10-20200304Meli UrtadoAún no hay calificaciones
- Dieta Del Chorlito Doble Collar (Charadrius Falklandicus)Documento7 páginasDieta Del Chorlito Doble Collar (Charadrius Falklandicus)Esteban VegaAún no hay calificaciones
- Vizcarra 2008 Avifauna Humedales Ite PeruDocumento22 páginasVizcarra 2008 Avifauna Humedales Ite PeruMarcelo Mayorga RodríguezAún no hay calificaciones
- 124-Texto Del Artículo-1153-1-10-20200528Documento3 páginas124-Texto Del Artículo-1153-1-10-20200528Katherin RendonAún no hay calificaciones
- 631-Texto Del Artículo-680-7-10-20141211Documento16 páginas631-Texto Del Artículo-680-7-10-20141211sundancebagsAún no hay calificaciones
- HABITOS ALIMENTARIOS DEL LAGARTO Cnemidophorus LemDocumento13 páginasHABITOS ALIMENTARIOS DEL LAGARTO Cnemidophorus LemAlejandroLópezAún no hay calificaciones
- Asociaciones Ecológicas en La Comunidad de Peces de La Porción Occidental de La Costa de CampecheDocumento16 páginasAsociaciones Ecológicas en La Comunidad de Peces de La Porción Occidental de La Costa de CampecheErick BarriosAún no hay calificaciones
- Emerita SP en CubaDocumento10 páginasEmerita SP en CubaFer MartellAún no hay calificaciones
- 505 - Aves Acuáticas en Ecosistemas Playeros Del Parque Nacional Cahuita, Limón, Costa RicaDocumento15 páginas505 - Aves Acuáticas en Ecosistemas Playeros Del Parque Nacional Cahuita, Limón, Costa RicaAnonymous XXENB3eOAún no hay calificaciones
- Efectos de La Pesqueria Industriales de Palangre Sobre Las Aves MarinasDocumento6 páginasEfectos de La Pesqueria Industriales de Palangre Sobre Las Aves MarinasMarcimio Pethelo ManoPello ProvosteAún no hay calificaciones
- Avesmarinas PDFDocumento8 páginasAvesmarinas PDFYola PortilloAún no hay calificaciones
- Parametros Reprodutivos Podocnemis - Carvajal - 2011Documento10 páginasParametros Reprodutivos Podocnemis - Carvajal - 2011Tanita Tana Taner TananaAún no hay calificaciones
- Informes Sobre TortugasDocumento12 páginasInformes Sobre TortugasFernando Cabezas PraviaAún no hay calificaciones
- Monitoreo de Poblaciones y Condición de Salud de Aves Marinas y Lobos Marinos en Islas Del Norte Del Golfo de California, MéxicoDocumento15 páginasMonitoreo de Poblaciones y Condición de Salud de Aves Marinas y Lobos Marinos en Islas Del Norte Del Golfo de California, MéxicoTiffany RengifoAún no hay calificaciones
- Informe 47 (1) Art 5 PDFDocumento9 páginasInforme 47 (1) Art 5 PDFWilly Alejandro Hernandez ChinarroAún no hay calificaciones
- RIM 41 (2) Art 3Documento15 páginasRIM 41 (2) Art 3GabrielAún no hay calificaciones
- Informe La Anciana 12 01 10Documento46 páginasInforme La Anciana 12 01 10Raquel ChavarriaAún no hay calificaciones
- ArrecifesDocumento4 páginasArrecifesGael Garcia LpzAún no hay calificaciones
- Traduccion ArticuloDocumento13 páginasTraduccion Articuloluis medina “el medis”Aún no hay calificaciones
- Arrecifes Coralinos Isla FuerteDocumento7 páginasArrecifes Coralinos Isla Fuertegustavo AraqueAún no hay calificaciones
- Uprevistas, 3Documento16 páginasUprevistas, 3Adrian CodesAún no hay calificaciones
- Dieta Del Biguá (Phalacrocorax Olivaceus) Durante La Primavera en El Estuario de Bahía Blanca, Buenos Aires, ArgentinaDocumento6 páginasDieta Del Biguá (Phalacrocorax Olivaceus) Durante La Primavera en El Estuario de Bahía Blanca, Buenos Aires, ArgentinaTalita BackhausAún no hay calificaciones
- Etnoictiología Trófica Tikuna de Airú A Yutá, en La Amazonia Colombiana-Junio-17Documento5 páginasEtnoictiología Trófica Tikuna de Airú A Yutá, en La Amazonia Colombiana-Junio-17Edgar Francisco Prieto PiraquiveAún no hay calificaciones
- Patrones de Presencia y Abundancia de Aves Terrestres en La Isla SaliacaDocumento8 páginasPatrones de Presencia y Abundancia de Aves Terrestres en La Isla Saliacaa21010007Aún no hay calificaciones
- Biodiv118art3c PDFDocumento5 páginasBiodiv118art3c PDFDiana BarrónAún no hay calificaciones
- 7334 PDFDocumento5 páginas7334 PDFKarelyy RmAún no hay calificaciones
- Tésis Elí García-PadillaDocumento40 páginasTésis Elí García-Padillaeligarcia_18Aún no hay calificaciones
- Tortugas: Fichas de Los TaxonesDocumento14 páginasTortugas: Fichas de Los Taxones86 leonelliniAún no hay calificaciones
- Pepinos de MarDocumento9 páginasPepinos de MarJaime EspinoAún no hay calificaciones
- Tea Final ComunidadesDocumento23 páginasTea Final ComunidadesJulia Elena Sánchez LópezAún no hay calificaciones
- Capitulo 9 Pesquerias Mexicanas de Tiburon en El Oceano Pacifico - CompressDocumento32 páginasCapitulo 9 Pesquerias Mexicanas de Tiburon en El Oceano Pacifico - CompressGabriel RodríguezAún no hay calificaciones
- Informe 44 (1) - 10Documento21 páginasInforme 44 (1) - 10TITO SILVA CHIROQUEAún no hay calificaciones
- Estructura de La Comunidad de Peces en Cuatro Estuarios Del Pacífico MexicanoDocumento13 páginasEstructura de La Comunidad de Peces en Cuatro Estuarios Del Pacífico MexicanoEdgar SandovalAún no hay calificaciones
- Manati Especie SombrillaDocumento11 páginasManati Especie SombrillaEnrique ZadomenAún no hay calificaciones
- Humedales de Ventanilla Acuario Natural PDFDocumento55 páginasHumedales de Ventanilla Acuario Natural PDFKarla RicaldiiAún no hay calificaciones
- Cacerolita de Mar ArtrevDocumento6 páginasCacerolita de Mar ArtrevSebastián Barragán100% (1)
- Bivalvos. Nota CientificaDocumento10 páginasBivalvos. Nota CientificaGloria Elena Gutierrez MoralesAún no hay calificaciones
- Articles-64772 DocumentoDocumento27 páginasArticles-64772 Documentoluis roberto aguilera tapiaAún no hay calificaciones
- Datos Ballena Azul PDFDocumento5 páginasDatos Ballena Azul PDFEduardo GutierrezAún no hay calificaciones
- Nuevos Registros de Moluscos Marinos de El Salvador, América CentralDocumento8 páginasNuevos Registros de Moluscos Marinos de El Salvador, América CentralCarlos RiveraAún no hay calificaciones
- Captura Incidental de La Raya Hypanus Americanus (Elasmobranchii) en La Pesca de Arrastre de Camarón en La Región Suroriental de CubaDocumento6 páginasCaptura Incidental de La Raya Hypanus Americanus (Elasmobranchii) en La Pesca de Arrastre de Camarón en La Región Suroriental de CubaKatherin RendonAún no hay calificaciones
- Lira, C. Et Al. 2012,. Crustáceos Decápodos Bentónicos Litorales de La Isla de La Tortuga. I. - Familia Porcellanidae Haworth, 1825Documento15 páginasLira, C. Et Al. 2012,. Crustáceos Decápodos Bentónicos Litorales de La Isla de La Tortuga. I. - Familia Porcellanidae Haworth, 1825Duver FernandezAún no hay calificaciones
- Cubomed PVRDocumento3 páginasCubomed PVRmax minAún no hay calificaciones
- RECOMENDACIONES PARA LA CONSERVACION EFECTIVA DE Lontra Felina EN PERUDocumento47 páginasRECOMENDACIONES PARA LA CONSERVACION EFECTIVA DE Lontra Felina EN PERUMiguel90Aún no hay calificaciones
- Cladoceros Colombia PDFDocumento42 páginasCladoceros Colombia PDFroraysi_cm01Aún no hay calificaciones
- Planificacion Espacil MarinaDocumento15 páginasPlanificacion Espacil MarinaAlejandro CalloniAún no hay calificaciones
- Tesoro-Ecologico-En-Riesgo ManglarDocumento5 páginasTesoro-Ecologico-En-Riesgo ManglarHector OlguinAún no hay calificaciones
- 806 2016 Chura Apaza Ki Faci Biologia MicrobiologiaDocumento176 páginas806 2016 Chura Apaza Ki Faci Biologia MicrobiologiaVanne BMAún no hay calificaciones
- T Roman Tesis Trichoderma 2019 Junio FinalDocumento46 páginasT Roman Tesis Trichoderma 2019 Junio FinalVanne BMAún no hay calificaciones
- GESTION MANEJO CONSERVACION ANPsDocumento64 páginasGESTION MANEJO CONSERVACION ANPsVanne BMAún no hay calificaciones
- Gaviotin GeneralidadesDocumento4 páginasGaviotin GeneralidadesVanne BMAún no hay calificaciones
- Trabajo de Responsabilidad SocialDocumento19 páginasTrabajo de Responsabilidad SocialRosario Inés Sánchez CamonesAún no hay calificaciones
- AntialergicosDocumento34 páginasAntialergicosMauricioAún no hay calificaciones
- Compuestos Bioactivos - Yulisa MorenoDocumento14 páginasCompuestos Bioactivos - Yulisa MorenoMina Lawliet0% (1)
- Aparato Reproductor MasculinoDocumento1 páginaAparato Reproductor MasculinoArturo AhumadaAún no hay calificaciones
- Diseño de Una Planta para La Producción de Compost en La Zona Norte Del Estado AnzoateguiDocumento224 páginasDiseño de Una Planta para La Producción de Compost en La Zona Norte Del Estado AnzoateguiJesus FrancoAún no hay calificaciones
- Concentracion GravimetricaDocumento10 páginasConcentracion GravimetricaAlí Naveros JuncoAún no hay calificaciones
- Places OrquitisDocumento12 páginasPlaces OrquitisElizabeth ÁlvarezAún no hay calificaciones
- Distalización de Molares Superiores Con Péndulo de Hilgers en Paciente Con Síndrome de Di GeorgeDocumento37 páginasDistalización de Molares Superiores Con Péndulo de Hilgers en Paciente Con Síndrome de Di GeorgeJavi Jaramillo100% (1)
- Martínez (2015) Eval Calidad Microbio Pescado Crudo Comercializado Puerto PDFDocumento223 páginasMartínez (2015) Eval Calidad Microbio Pescado Crudo Comercializado Puerto PDFNatalia SHAún no hay calificaciones
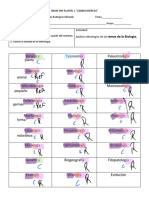
- Ramas de La Biologia 2022-23Documento3 páginasRamas de La Biologia 2022-23Iker MonroyAún no hay calificaciones
- Yoga Nidra - Script en EspañolDocumento4 páginasYoga Nidra - Script en EspañolAlex Macias75% (4)
- Ejercicios de GeneticaDocumento7 páginasEjercicios de GeneticaNormaAún no hay calificaciones
- Individual A Las 6Documento3 páginasIndividual A Las 6Brenda Cupitay HuamaniAún no hay calificaciones
- Codex Alimentarius Sacha Inchi 2.021Documento14 páginasCodex Alimentarius Sacha Inchi 2.021Oscar PeñaAún no hay calificaciones
- AsfixiaDocumento50 páginasAsfixiaDivisay VenenciaAún no hay calificaciones
- Ramas de La SociedadDocumento10 páginasRamas de La SociedadEduardoNezzTejadaAún no hay calificaciones
- Exposicion Salud OcupacionalDocumento6 páginasExposicion Salud OcupacionalSOFIA VALENTINA SUAREZ TORRESAún no hay calificaciones
- Síndrome NefróticoDocumento50 páginasSíndrome NefróticoMariajo RochaAún no hay calificaciones
- Diapositivas de Psicologia 1Documento12 páginasDiapositivas de Psicologia 1Calidad Santo TomasAún no hay calificaciones
- Portafolio PinzasDocumento11 páginasPortafolio PinzasCriss VillegasAún no hay calificaciones
- Estrategias CatalíticasDocumento20 páginasEstrategias CatalíticasjoselynAún no hay calificaciones
- Taller de Repaso7Documento6 páginasTaller de Repaso7SugeyAún no hay calificaciones
- Bioelectricidad, Los Secretos de La Regeneración Humana - Cinco Noticias - IMPORTANTEDocumento6 páginasBioelectricidad, Los Secretos de La Regeneración Humana - Cinco Noticias - IMPORTANTEAndrosamuel GonzalezvalenzuelaAún no hay calificaciones
- Escala de Inadaptación (Estrés Postraumático)Documento2 páginasEscala de Inadaptación (Estrés Postraumático)Nuria de Lamo100% (2)
- Trabajo de Biologia IIDocumento5 páginasTrabajo de Biologia IIJosé Nerio FloresAún no hay calificaciones
- Ejerciciospropuestos de Enlaces QuimicosDocumento4 páginasEjerciciospropuestos de Enlaces QuimicosLeonardo Condori Mamani0% (1)
- Orto Implantes - 1 Clase. 2Documento34 páginasOrto Implantes - 1 Clase. 2ana100% (1)
- Origenes R Shapiro Biblioteca Cientifica Salvat 054 1994 OCRDocumento165 páginasOrigenes R Shapiro Biblioteca Cientifica Salvat 054 1994 OCREnrique Luis CoronelAún no hay calificaciones
- Manual de Bioseguridad en El Laboratorio Cuarta EdiciónDocumento125 páginasManual de Bioseguridad en El Laboratorio Cuarta EdiciónbettyAún no hay calificaciones
- Informe Técnico de Impactos Causados A Nivel de Producción de Alimentos en El Municipio de BarbacoasDocumento10 páginasInforme Técnico de Impactos Causados A Nivel de Producción de Alimentos en El Municipio de BarbacoasHernan LopezAún no hay calificaciones