También podría gustarte
- NeurogliaDocumento37 páginasNeurogliaMaria camila Torres jerezAún no hay calificaciones
- Abriendo la caja negra: Una historia de la neurocienciaDe EverandAbriendo la caja negra: Una historia de la neurocienciaAún no hay calificaciones
- NEURONADocumento3 páginasNEURONAleonardo david pinzónAún no hay calificaciones
- A cada cual su cerebro: Plasticidad neuronal e inconscienteDe EverandA cada cual su cerebro: Plasticidad neuronal e inconscienteAún no hay calificaciones
- Ramón y Cajal y GolgiDocumento7 páginasRamón y Cajal y GolgiCristian Muñoz BustosAún no hay calificaciones
- Duelo de Titanes Camillo Golgi y Santiago Ramón y CajalDocumento3 páginasDuelo de Titanes Camillo Golgi y Santiago Ramón y CajalCintia AyllónAún no hay calificaciones
- Resumen de Santiago Ramón y Cajal y Las Aportaciones Al Campo de Las NeurocienciasDocumento5 páginasResumen de Santiago Ramón y Cajal y Las Aportaciones Al Campo de Las NeurocienciasZFI aduanasAún no hay calificaciones
- NeurocienciaDocumento5 páginasNeurocienciaMartín Lucas Chavarría ReynagaAún no hay calificaciones
- La Glía y CélulasDocumento20 páginasLa Glía y CélulasVivian ItzelAún no hay calificaciones
- NeurobiologíaDocumento8 páginasNeurobiologíaSylwia AnnaAún no hay calificaciones
- NeurobiolgiaDocumento10 páginasNeurobiolgiaLeon Elorza BeatrizAún no hay calificaciones
- Historia de Neurociencias - Bases BiológicasDocumento9 páginasHistoria de Neurociencias - Bases Biológicasnicole yarlaqueAún no hay calificaciones
- Epistemología de La NEUROPSICOLOGÍADocumento10 páginasEpistemología de La NEUROPSICOLOGÍAAngel de leonAún no hay calificaciones
- Ramon y Cajal y La Neurociencia Del SigloDocumento5 páginasRamon y Cajal y La Neurociencia Del Sigloegcc5Aún no hay calificaciones
- Ramon Cajal y La Neurociencia en El Siglo XXIDocumento5 páginasRamon Cajal y La Neurociencia en El Siglo XXIxlichun23Aún no hay calificaciones
- Santiago Ramón y Teoria CelularDocumento3 páginasSantiago Ramón y Teoria CelularMisaelAún no hay calificaciones
- NEUROPSICOLOGÍADocumento8 páginasNEUROPSICOLOGÍAAdriana Gil Echeverry100% (1)
- Cap I - Introduccion A La NeurocienciaDocumento11 páginasCap I - Introduccion A La NeurocienciaClara EddyAún no hay calificaciones
- AFTmarzo2008 Historia NeurotransDocumento7 páginasAFTmarzo2008 Historia NeurotransFarmaFM100% (2)
- Historia de La NeurocienciaDocumento4 páginasHistoria de La Neurocienciagregomar100% (1)
- Células GlialesDocumento4 páginasCélulas GlialesMaria Reyna Guerrero BalderasAún no hay calificaciones
- HISTORIADocumento2 páginasHISTORIAZulmith NayeliAún no hay calificaciones
- Eric KandelDocumento11 páginasEric KandelGiancarlo Moya100% (3)
- Doctrina de La NeuronaDocumento2 páginasDoctrina de La NeuronaGiovannaAún no hay calificaciones
- Las Microglías.-Actúan Durante Una Inflamación yDocumento2 páginasLas Microglías.-Actúan Durante Una Inflamación yEdy MonrroyAún no hay calificaciones
- Historia NeurologíaDocumento5 páginasHistoria NeurologíaDenis MoraAún no hay calificaciones
- Curso: "Año Del Fortalecimiento de La Soberanía Nacional"Documento6 páginasCurso: "Año Del Fortalecimiento de La Soberanía Nacional"DINA MARISOL PARIAPAZA MACHACAAún no hay calificaciones
- Cerebro y MenteDocumento3 páginasCerebro y MenteRodolfo ChuquimbalquiAún no hay calificaciones
- EnsayoDocumento6 páginasEnsayoMauricio PérezAún no hay calificaciones
- Origen de La NeurocienciaDocumento7 páginasOrigen de La NeurocienciaVictor Díaz AlarconAún no hay calificaciones
- Fundamentos de NeuroanatomiaDocumento8 páginasFundamentos de NeuroanatomiaCoral LunaAún no hay calificaciones
- NEUROGLÍADocumento11 páginasNEUROGLÍAJimmy RobertoAún no hay calificaciones
- Síntesis de La Neurociencia y Su Historia Coralia Ramírez Ramírez 19-EPST-1-131 PDFDocumento5 páginasSíntesis de La Neurociencia y Su Historia Coralia Ramírez Ramírez 19-EPST-1-131 PDFNorelys Germosen AcevedoAún no hay calificaciones
- Antecedentes y Perspectivas de La Neuropsicología ActualDocumento10 páginasAntecedentes y Perspectivas de La Neuropsicología ActualIván FigueroaAún no hay calificaciones
- BiomoleculasDocumento4 páginasBiomoleculasjuanAún no hay calificaciones
- Clase 02Documento14 páginasClase 02JesúsCoyacAún no hay calificaciones
- El Cerebro y El ComportamientoDocumento17 páginasEl Cerebro y El ComportamientoKatherin castellanosAún no hay calificaciones
- Neuroglias Martin RodriguezDocumento5 páginasNeuroglias Martin RodriguezMartinRodriguezBuelaAún no hay calificaciones
- Neurociencia CognitivaDocumento4 páginasNeurociencia CognitivaalbertoAún no hay calificaciones
- Estructura Interna de Las NeuronasDocumento33 páginasEstructura Interna de Las NeuronasNina Ninón100% (3)
- Neurociencia Cognitiva PDFDocumento4 páginasNeurociencia Cognitiva PDFMayra HerreraAún no hay calificaciones
- Camillo GolgiDocumento6 páginasCamillo GolgiInma MaldonadoAún no hay calificaciones
- Modelos en PsicopatologíaDocumento37 páginasModelos en PsicopatologíaDiana RodAún no hay calificaciones
- Tarea 6 de Teoria de La Psicologia ActualesDocumento5 páginasTarea 6 de Teoria de La Psicologia ActualesbriannaAún no hay calificaciones
- Linea de Tiempo de NeurocienciasDocumento3 páginasLinea de Tiempo de NeurocienciasIrviana ÁlvarezAún no hay calificaciones
- C2 Historia de La NeurocienciaDocumento5 páginasC2 Historia de La NeurocienciaPAULA ANDREA NUÑEZ ORTIZAún no hay calificaciones
- Relatos Desde Los Dos Lados Del Cerebro-Sergio TamayoDocumento36 páginasRelatos Desde Los Dos Lados Del Cerebro-Sergio TamayoPattiRivera100% (1)
- Relatos Desde Los Dos Lados Del Cerebro-Sergio TamayoDocumento41 páginasRelatos Desde Los Dos Lados Del Cerebro-Sergio TamayoPattiRivera0% (1)
- El Universo InteriorDocumento9 páginasEl Universo InterioreduardoAún no hay calificaciones
- Santiago Ramón y Cajal Nació El 1 de Mayo de 1852 en Petilla de AragónDocumento6 páginasSantiago Ramón y Cajal Nació El 1 de Mayo de 1852 en Petilla de AragónFANNYCUELLARAún no hay calificaciones
- CISNEROS GARCIAS M - La Historia de Las Neurociencias y Los MetodosDocumento3 páginasCISNEROS GARCIAS M - La Historia de Las Neurociencias y Los MetodosFernanda CisnerosAún no hay calificaciones
- Reseña Histórica de La Histología y Del MicroscopioDocumento3 páginasReseña Histórica de La Histología y Del MicroscopioLIDIA MERCEDES MARTINEZ CRUZAún no hay calificaciones
- Bases BiologicasDocumento8 páginasBases BiologicasAlessandra TesenAún no hay calificaciones
- Cap2. Neurociencia y AprendizajeDocumento19 páginasCap2. Neurociencia y AprendizajePaola QuisbertAún no hay calificaciones
- 2 Cap. Neurociencia PUCP 1Documento10 páginas2 Cap. Neurociencia PUCP 1Lucelia Ramirez MillaAún no hay calificaciones
- Tejido NerrviosoDocumento17 páginasTejido NerrviosoLiz AbeledoAún no hay calificaciones
- Neurociencia Del Siglo XXI Ramon y CajalDocumento5 páginasNeurociencia Del Siglo XXI Ramon y CajalPaolo MoralesAún no hay calificaciones
- Linea Del TiempoDocumento3 páginasLinea Del TiempoVero BernalAún no hay calificaciones
- Cerebro Conducta....... Enfoques Que Describen La Relacion Cerebro-ConductaDocumento7 páginasCerebro Conducta....... Enfoques Que Describen La Relacion Cerebro-ConductaJohanna Ponce0% (1)
- Segundo Parcial Histologà A Neuro SRDocumento9 páginasSegundo Parcial Histologà A Neuro SRjoão dornellesAún no hay calificaciones
- Parcial 2 SRDocumento17 páginasParcial 2 SRjoão dornellesAún no hay calificaciones
- Preguntas Del Resumen HistologíaDocumento1 páginaPreguntas Del Resumen Histologíajoão dornellesAún no hay calificaciones
- Segundo Parcial Histologà A Neuro SRDocumento9 páginasSegundo Parcial Histologà A Neuro SRjoão dornellesAún no hay calificaciones
- Parcial 2 SRDocumento17 páginasParcial 2 SRjoão dornellesAún no hay calificaciones
- HNCR - Parcial 13.11.2021 SRDocumento7 páginasHNCR - Parcial 13.11.2021 SRjoão dornellesAún no hay calificaciones
- HNCR - Parcial 13.11.2021 SRDocumento7 páginasHNCR - Parcial 13.11.2021 SRjoão dornellesAún no hay calificaciones
- Clases G2Documento1 páginaClases G2joão dornellesAún no hay calificaciones
- Uc 8 - Anexo Ii Histob 2020Documento2 páginasUc 8 - Anexo Ii Histob 2020joão dornellesAún no hay calificaciones
- UC 8 - Programa HISTOB 2020Documento4 páginasUC 8 - Programa HISTOB 2020joão dornellesAún no hay calificaciones
- Presentación 1Documento1 páginaPresentación 1joão dornellesAún no hay calificaciones
- Protocolo Prácticos Presenciales Anatomía 2022 en El Contexto de La Pandemia Covid 19Documento3 páginasProtocolo Prácticos Presenciales Anatomía 2022 en El Contexto de La Pandemia Covid 19joão dornellesAún no hay calificaciones
- Cronograma Anatomía Clínica 2021Documento2 páginasCronograma Anatomía Clínica 2021joão dornellesAún no hay calificaciones
- Cronograma Clases Teóricas en You Tube Anatomía Clínica 2022Documento10 páginasCronograma Clases Teóricas en You Tube Anatomía Clínica 2022joão dornellesAún no hay calificaciones
- Resumen BCMDocumento171 páginasResumen BCMAgustín BiurraAún no hay calificaciones
- EeeeeDocumento1 páginaEeeeejoão dornellesAún no hay calificaciones
- Glucólisis: (Gluco Azúcar Lisis Destrucción o Rotura)Documento85 páginasGlucólisis: (Gluco Azúcar Lisis Destrucción o Rotura)joão dornellesAún no hay calificaciones
- Curva de Crecimiento - Biofísica.Documento3 páginasCurva de Crecimiento - Biofísica.joão dornellesAún no hay calificaciones
- MSP - HCU - Certificadovacunacion10010571 GARCIA FATIMADocumento2 páginasMSP - HCU - Certificadovacunacion10010571 GARCIA FATIMAgeorgechuchuca23100% (1)
- Enferpedia - Técnicas y Procedimientos de EnfermeríaDocumento41 páginasEnferpedia - Técnicas y Procedimientos de EnfermeríaOscar Sanchez100% (1)
- Manuscrito Tesis II-mariela BustamanteDocumento72 páginasManuscrito Tesis II-mariela Bustamantecamila andrea lizana osorioAún no hay calificaciones
- AntihipertensivosDocumento15 páginasAntihipertensivosArabeela Sanchez QuispeAún no hay calificaciones
- Proteomica y Farmacia. TraduccionDocumento9 páginasProteomica y Farmacia. TraduccionAlejandra NeisaAún no hay calificaciones
- PRUNAPEDocumento12 páginasPRUNAPESteff WelchAún no hay calificaciones
- OBSTRUCCION INTESTINALmbhjDocumento5 páginasOBSTRUCCION INTESTINALmbhjAlejandra Rodríguez SalgadoAún no hay calificaciones
- Trabajo de Psicologia SocialDocumento11 páginasTrabajo de Psicologia SocialDaymoon zombieAún no hay calificaciones
- Lista de Ejercicios 03Documento2 páginasLista de Ejercicios 03Mario Broster GelatinaAún no hay calificaciones
- Taller Equilibrio Acido Basico.Documento6 páginasTaller Equilibrio Acido Basico.Aady Meli FernándezAún no hay calificaciones
- Desarrollo de Los Niños en La Edad EscolarDocumento6 páginasDesarrollo de Los Niños en La Edad EscolarLuchy CornieleAún no hay calificaciones
- 11.conceptos Básicos de La Ventilación Mecánica PDFDocumento30 páginas11.conceptos Básicos de La Ventilación Mecánica PDFMARIA CAMILA GUZMAN CANARIA100% (1)
- Casa BlancaDocumento11 páginasCasa BlancaAlejandro Alvarado CotrinaAún no hay calificaciones
- Document PDFDocumento9 páginasDocument PDFEFRANOR CRACK 2Aún no hay calificaciones
- Semiología de RodillaDocumento31 páginasSemiología de RodillaCarla100% (2)
- Cálculos y DilucionesDocumento6 páginasCálculos y DilucionesSara Pamela LondoñoAún no hay calificaciones
- Unidad 7 - Gestión de Muestras Biológicas IIDocumento6 páginasUnidad 7 - Gestión de Muestras Biológicas IIPAULA UBEDA LOPEZAún no hay calificaciones
- Ficha Práctica 03. Asegurar Que Las Niñas y Niños Reciban Los Cuidados Necesarios para Una Buena Salud Eliana 2Documento5 páginasFicha Práctica 03. Asegurar Que Las Niñas y Niños Reciban Los Cuidados Necesarios para Una Buena Salud Eliana 2Beranis Torres100% (1)
- Actividad Fisica en La AdolescenciaDocumento14 páginasActividad Fisica en La AdolescenciaRafael LinaresAún no hay calificaciones
- Examenes FARMADocumento54 páginasExamenes FARMACarmen Llorente100% (1)
- SG-PR-09 Exámenes MedicosDocumento6 páginasSG-PR-09 Exámenes MedicosPaola Andrea Sepulveda VargasAún no hay calificaciones
- PIROPLASMOSISDocumento9 páginasPIROPLASMOSISDIANA VALLEJOS RODRIGUEZAún no hay calificaciones
- Actividad Eje N.2 Metodologia de La InvestigacionDocumento6 páginasActividad Eje N.2 Metodologia de La InvestigacionLisbeth LópezAún no hay calificaciones
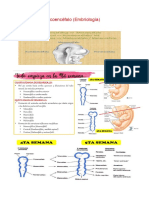
- Semana 2 - Troncoencéfalo (Embriología)Documento11 páginasSemana 2 - Troncoencéfalo (Embriología)mariano arroyo ojedaAún no hay calificaciones
- ShockDocumento8 páginasShockKaren Alejandra ParodiiAún no hay calificaciones
- ColesterolDocumento2 páginasColesterolRicardo Ortiz Ramos100% (2)
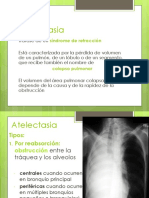
- Atelectasias DerrameDocumento72 páginasAtelectasias DerrameJose Luis Foronda Rios100% (1)
- Mieloma MultipleDocumento15 páginasMieloma MultipleNaye Recalde ErazoAún no hay calificaciones
- Mof Sedes 2018 - 0Documento121 páginasMof Sedes 2018 - 0Gonzalo Coaquira100% (1)
- UntitledDocumento6 páginasUntitledEstefania GonzálezAún no hay calificaciones
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- Cómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeDe EverandCómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeCalificación: 4 de 5 estrellas4/5 (81)
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- Autismo: guía para padres sobre el trastorno del espectro autista En españolDe EverandAutismo: guía para padres sobre el trastorno del espectro autista En españolCalificación: 4.5 de 5 estrellas4.5/5 (4)
- Psicología y trastornos de los niños en la edad evolutiva: Qué son y cómo funcionanDe EverandPsicología y trastornos de los niños en la edad evolutiva: Qué son y cómo funcionanCalificación: 5 de 5 estrellas5/5 (1)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Cómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasDe EverandCómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasCalificación: 4.5 de 5 estrellas4.5/5 (12)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Neurociencia para vencer la depresión: La esprial ascendenteDe EverandNeurociencia para vencer la depresión: La esprial ascendenteCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Dinámicas familiares a través de la vida de José: La túnica del PadreDe EverandDinámicas familiares a través de la vida de José: La túnica del PadreCalificación: 5 de 5 estrellas5/5 (2)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Neuropsicología: Los fundamentos de la materiaDe EverandNeuropsicología: Los fundamentos de la materiaCalificación: 5 de 5 estrellas5/5 (1)
- Terapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónDe EverandTerapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónCalificación: 5 de 5 estrellas5/5 (3)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- Bioneuroemoción: Un método para el bienestar emocionalDe EverandBioneuroemoción: Un método para el bienestar emocionalCalificación: 5 de 5 estrellas5/5 (4)
- Psicópatas seriales: Un recorrido por su oscura e inquietante naturalezaDe EverandPsicópatas seriales: Un recorrido por su oscura e inquietante naturalezaCalificación: 4 de 5 estrellas4/5 (3)
- Nuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosDe EverandNuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosCalificación: 3.5 de 5 estrellas3.5/5 (6)
- El poder de la presencia: Cómo la presencia de los padres moldea el cerebro de los hijos y configura las personas que llegarán a serDe EverandEl poder de la presencia: Cómo la presencia de los padres moldea el cerebro de los hijos y configura las personas que llegarán a serCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Agilidad emocional: Rompe tus bloqueos, abraza el cambio y triunfa en el trabajo y en la vdaDe EverandAgilidad emocional: Rompe tus bloqueos, abraza el cambio y triunfa en el trabajo y en la vdaCalificación: 4.5 de 5 estrellas4.5/5 (34)
- La conciencia en el cerebro: Descifrando el enigma de cómo el cerebro elabora nuestros pensamientosDe EverandLa conciencia en el cerebro: Descifrando el enigma de cómo el cerebro elabora nuestros pensamientosCalificación: 2.5 de 5 estrellas2.5/5 (3)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- Manual de psicoterapia emocional sistémicaDe EverandManual de psicoterapia emocional sistémicaCalificación: 4.5 de 5 estrellas4.5/5 (7)
- Mindfulness: Cómo hacer menos y ser más conscientes puede ayudarnos a vivir mejor en este mundo estresadoDe EverandMindfulness: Cómo hacer menos y ser más conscientes puede ayudarnos a vivir mejor en este mundo estresadoCalificación: 4.5 de 5 estrellas4.5/5 (9)
- PNL: Programación neurolingüística: Una guía práctica y sencilla para iniciarse en la programación neurolingüísticaDe EverandPNL: Programación neurolingüística: Una guía práctica y sencilla para iniciarse en la programación neurolingüísticaCalificación: 4 de 5 estrellas4/5 (65)