También podría gustarte
- El Origen de La PluricelularidadDocumento1 páginaEl Origen de La PluricelularidadJuanjo GayossoAún no hay calificaciones
- Reproduccion de Los Seres VivosDocumento15 páginasReproduccion de Los Seres VivosEdixon CastiblancoAún no hay calificaciones
- Transporte de Sustancias (Organismos Vivos)Documento17 páginasTransporte de Sustancias (Organismos Vivos)Edgar Gerardo Borrego PeñaAún no hay calificaciones
- La NutricionDocumento2 páginasLa NutricionWaldemar Muñoz RuizAún no hay calificaciones
- Reproducción Sexual en PlantasDocumento2 páginasReproducción Sexual en PlantasJHONNY100% (1)
- Monsta XDocumento8 páginasMonsta XLaura OtalvaroAún no hay calificaciones
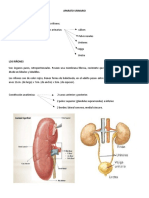
- Sistema Excretor: ¿Qué Es?: ¿Cuáles Son Las Estructuras Del Sistema Excretor?Documento12 páginasSistema Excretor: ¿Qué Es?: ¿Cuáles Son Las Estructuras Del Sistema Excretor?YOELAún no hay calificaciones
- AnimalesDocumento15 páginasAnimalesMarjorie Lizeth Tineo DíazAún no hay calificaciones
- Diferencias Estructurales de La Celula Animal y VegetalDocumento12 páginasDiferencias Estructurales de La Celula Animal y VegetalCarolina MtzAún no hay calificaciones
- Guía de Actividades y Rúbrica Evaluación - Tarea 3 - Metabolismo Catabolismo y AnabolismoDocumento14 páginasGuía de Actividades y Rúbrica Evaluación - Tarea 3 - Metabolismo Catabolismo y AnabolismoTATIANAAún no hay calificaciones
- Marco TeóricoDocumento9 páginasMarco TeóricoMariana Alviso100% (1)
- Capitulo 11 Sistema MuscularDocumento1 páginaCapitulo 11 Sistema MuscularPercySalazarAún no hay calificaciones
- Teoria de La EvolucionDocumento9 páginasTeoria de La EvolucionMarianaFlores0% (1)
- ARACNIDOSDocumento5 páginasARACNIDOSElena Carhuatanta HernándezAún no hay calificaciones
- Aparato Reproductor en Aves HembraDocumento4 páginasAparato Reproductor en Aves Hembraanthonella.reyesAún no hay calificaciones
- Sistema Reproductor Humano-1Documento4 páginasSistema Reproductor Humano-1asleitAún no hay calificaciones
- Sistema Renal o UrinarioDocumento2 páginasSistema Renal o UrinarioLizette CalzadaAún no hay calificaciones
- Sistema ReproductorDocumento21 páginasSistema ReproductorGy William Em CyAún no hay calificaciones
- 465 2013 08 22 C3Trematodos PDFDocumento5 páginas465 2013 08 22 C3Trematodos PDFLaura BernalAún no hay calificaciones
- Practica-1 Origen de La VidaDocumento9 páginasPractica-1 Origen de La VidaLídia BatistaAún no hay calificaciones
- Espermatogénesis y OvogenesisDocumento25 páginasEspermatogénesis y OvogenesisIsrael Silva SaucedoAún no hay calificaciones
- ApéndiceDocumento2 páginasApéndiceYesicaAún no hay calificaciones
- Desarrollo EmbrionarioDocumento17 páginasDesarrollo EmbrionarioJosue Coronado O. CndAún no hay calificaciones
- La-Reproducción I - PREDocumento5 páginasLa-Reproducción I - PREAlex CadilloAún no hay calificaciones
- La Teoría Celular y Sus PostulacionesDocumento3 páginasLa Teoría Celular y Sus PostulacionesAndy CuadradoAún no hay calificaciones
- Zoologia de PlatelmintosDocumento17 páginasZoologia de PlatelmintosJohn VelascoAún no hay calificaciones
- Sistema Respiratorio de Los AnfibiosDocumento5 páginasSistema Respiratorio de Los AnfibiosCarlos Rodriguez PantojaAún no hay calificaciones
- 01 Lo Vivo Difiere de Lo InerteDocumento2 páginas01 Lo Vivo Difiere de Lo InerteLucio RobertAún no hay calificaciones
- Locomoción en Seres VivosDocumento13 páginasLocomoción en Seres VivosSonia GuerreroAún no hay calificaciones
- Informe de Taxonomia de La Esponjas Marinas.Documento9 páginasInforme de Taxonomia de La Esponjas Marinas.Nicole Alexandra Vargas Rodriguez100% (1)
- Las Funciones de Nutrición en Los AnimalesDocumento5 páginasLas Funciones de Nutrición en Los Animalesmariyurmi garciaAún no hay calificaciones
- Marco TeóricoDocumento4 páginasMarco TeóricoMarlenny GonzalezAún no hay calificaciones
- INFORME 1 ProtozooariosDocumento11 páginasINFORME 1 ProtozooariosMarcela MosqueraAún no hay calificaciones
- Funciones Vitales de Los Seres VivosDocumento2 páginasFunciones Vitales de Los Seres VivosRenzo Iván Marín DávalosAún no hay calificaciones
- 59879583.trabajo Práctico Celulas - 2012Documento3 páginas59879583.trabajo Práctico Celulas - 2012Mariela SosaAún no hay calificaciones
- Funciones Del Aparato Reproductor MasculinoDocumento2 páginasFunciones Del Aparato Reproductor MasculinoKevin VelasquezAún no hay calificaciones
- Guía 9 Grupos Sanguíneos y RHDocumento3 páginasGuía 9 Grupos Sanguíneos y RHJUAN PABLO HOYOS TABARESAún no hay calificaciones
- Proceso de FecundaciónDocumento1 páginaProceso de FecundaciónSantiago Rguez OrozcoAún no hay calificaciones
- 01.que Es La Vida - CURTISDocumento3 páginas01.que Es La Vida - CURTISmadeAún no hay calificaciones
- Principales Características de Los Reinos BiologicosDocumento16 páginasPrincipales Características de Los Reinos BiologicosJairo A. Rivera0% (2)
- Diferencias en El Sistema Digestivo de Los Animales Vertebrados 5 Grado - 1Documento1 páginaDiferencias en El Sistema Digestivo de Los Animales Vertebrados 5 Grado - 1GENESIS NIKOL MONCADA ORTIZAún no hay calificaciones
- Quien Define El Sexo Del BebeDocumento2 páginasQuien Define El Sexo Del Bebegrest2100% (1)
- Guia N.13circulacion en Vertebrados y El HombreDocumento6 páginasGuia N.13circulacion en Vertebrados y El Hombrejuancik7215Aún no hay calificaciones
- Tema 10 Parte 2 Ciclo Desarrollo PlantasDocumento31 páginasTema 10 Parte 2 Ciclo Desarrollo PlantasBeatriz García CanoAún no hay calificaciones
- Características Generales Del Reino ProtistaDocumento6 páginasCaracterísticas Generales Del Reino ProtistaArq Adolfo Sánchez100% (1)
- Características de Los Seres VivosDocumento75 páginasCaracterísticas de Los Seres Vivosangie orizola caychoAún no hay calificaciones
- Aparato Reproductor Femenino y MasculinoDocumento2 páginasAparato Reproductor Femenino y MasculinoYuvilis RojasAún no hay calificaciones
- El Reino VegetalDocumento5 páginasEl Reino VegetalMaria Mendoza MendozaAún no hay calificaciones
- Actividad 38 FECUNDACIÓN Y EMBARAZODocumento6 páginasActividad 38 FECUNDACIÓN Y EMBARAZODaniela MelgarejoAún no hay calificaciones
- Informe de Laboratorio - Sistema Reprodutor FINALDocumento6 páginasInforme de Laboratorio - Sistema Reprodutor FINALEric Garcia RicardoAún no hay calificaciones
- Actividades Repaso Célula y Teoría EndosimbióticaDocumento2 páginasActividades Repaso Célula y Teoría EndosimbióticaAdriánSotoCuevasAún no hay calificaciones
- DesarrolloDocumento5 páginasDesarrolloMiguel PulidoAún no hay calificaciones
- Erizo de Mar LaboratorioDocumento9 páginasErizo de Mar LaboratorioValery Sue Barreda OrtizAún no hay calificaciones
- Guia de Estudio Sistema ExcretorDocumento12 páginasGuia de Estudio Sistema ExcretorAlexander León PuelloAún no hay calificaciones
- Que Es El MiscrocopioDocumento10 páginasQue Es El MiscrocopioValentina Pedrozo SuarezAún no hay calificaciones
- Aparato UrinarioDocumento9 páginasAparato Urinariomariana rodriguezAún no hay calificaciones
- Evaluacion Mensual de CtaDocumento2 páginasEvaluacion Mensual de CtaLedesma LópezAún no hay calificaciones
- Aparato UrinarioDocumento7 páginasAparato Urinariocarlos noriega diazAún no hay calificaciones
- Embriología de Las Vias UrinariasDocumento4 páginasEmbriología de Las Vias UrinariasPedro RodriguezAún no hay calificaciones
- Embriologia Del Sistema UrogenitalDocumento47 páginasEmbriologia Del Sistema UrogenitalHumberto Porras100% (1)
- CICATRIZACIÓNDocumento36 páginasCICATRIZACIÓNnahideli sarahi ochoa barajasAún no hay calificaciones
- Ciclo Ovarico y FecundacionDocumento4 páginasCiclo Ovarico y Fecundacionnahideli sarahi ochoa barajasAún no hay calificaciones
- ANTIMICOTICOSDocumento8 páginasANTIMICOTICOSnahideli sarahi ochoa barajasAún no hay calificaciones
- Anexos de La PielDocumento2 páginasAnexos de La Pielnahideli sarahi ochoa barajasAún no hay calificaciones
- Embarazo y LactanciaDocumento38 páginasEmbarazo y Lactancianahideli sarahi ochoa barajasAún no hay calificaciones
- AMINOACIDOSDocumento2 páginasAMINOACIDOSnahideli sarahi ochoa barajasAún no hay calificaciones
- Atencion de ConsultaDocumento8 páginasAtencion de Consultanahideli sarahi ochoa barajasAún no hay calificaciones
- 12) Módulo Plan de AcciónDocumento5 páginas12) Módulo Plan de Acciónnahideli sarahi ochoa barajasAún no hay calificaciones
- Anatomía (Cuello, M. Superior, Faringe)Documento37 páginasAnatomía (Cuello, M. Superior, Faringe)nahideli sarahi ochoa barajasAún no hay calificaciones
- Biosintesis y Degradacion de NucleotidosDocumento34 páginasBiosintesis y Degradacion de Nucleotidosnahideli sarahi ochoa barajasAún no hay calificaciones
- Sindrome de CushingDocumento2 páginasSindrome de Cushingnahideli sarahi ochoa barajasAún no hay calificaciones
- Exploracion Nariz, Oidos y GargantaDocumento1 páginaExploracion Nariz, Oidos y Gargantanahideli sarahi ochoa barajasAún no hay calificaciones
- Embarazo MúltipleDocumento14 páginasEmbarazo Múltiplediana carolina gomez sanchezAún no hay calificaciones
- Funcion de ReproduccionDocumento5 páginasFuncion de ReproduccionRugilup HernándezAún no hay calificaciones
- Emfermedades VenereasDocumento11 páginasEmfermedades VenereasCamilla Sánchez ArataAún no hay calificaciones
- Movimientos FetalesDocumento6 páginasMovimientos Fetalesanais_31nita0% (1)
- Métodos AnticonceptivosDocumento18 páginasMétodos AnticonceptivosWendy PulgarinAún no hay calificaciones
- 1 - Romper El Silencio Acerca de La MenstruacionDocumento7 páginas1 - Romper El Silencio Acerca de La MenstruacionVianey GarciaAún no hay calificaciones
- ETIMOLOGÍADocumento5 páginasETIMOLOGÍARocío FloresAún no hay calificaciones
- Lactancia Materna (Triptico) HSJD 2014Documento3 páginasLactancia Materna (Triptico) HSJD 2014Nutri JucarAún no hay calificaciones
- Sindrome de Patau Trisomia 13Documento1 páginaSindrome de Patau Trisomia 13LUIS ALBERTO LOPEZ DOMINGUEZAún no hay calificaciones
- Histeroscopia DiagnosticaDocumento5 páginasHisteroscopia DiagnosticaSofía SimpértigueAún no hay calificaciones
- INFERTILIDAD1Documento19 páginasINFERTILIDAD1MILENE ANGELICA NICOLE AMAYA BALDARRAGOAún no hay calificaciones
- La MenstruaciónDocumento3 páginasLa MenstruaciónLupitaAún no hay calificaciones
- Guia PracticaDocumento2 páginasGuia PracticaCopymundo CañarAún no hay calificaciones
- Antecedentes Materno InfantilDocumento4 páginasAntecedentes Materno InfantilFrancisco Javier León67% (3)
- Citología VaginalDocumento5 páginasCitología VaginalTATIANA MARILU SANCARRANCO ESTELAAún no hay calificaciones
- Tema 2 Las Funciones VitalesDocumento4 páginasTema 2 Las Funciones VitalesIreneAún no hay calificaciones
- EXAMEN EXTRAORDINARIO ALGAS 2021 DarioDocumento5 páginasEXAMEN EXTRAORDINARIO ALGAS 2021 DarioLinda PlasBurAún no hay calificaciones
- Arbol de Problemas AdryDocumento1 páginaArbol de Problemas AdryPaúl SalazarAún no hay calificaciones
- Me Corrí Con Mi Hermana.mDocumento10 páginasMe Corrí Con Mi Hermana.mRaul Alejandro Mujica SaraviaAún no hay calificaciones
- Revision Bibliografica de La Reproduccion Del Canis FamiliarisDocumento79 páginasRevision Bibliografica de La Reproduccion Del Canis FamiliarisGeovanni LunaAún no hay calificaciones
- AMENORREASDocumento30 páginasAMENORREASJennyMarcelaAristizabalAún no hay calificaciones
- Podcast Sobre Sex EducationDocumento4 páginasPodcast Sobre Sex EducationAna Cristina Guerrero MorenoAún no hay calificaciones
- Informe Semana AndinaDocumento8 páginasInforme Semana AndinaLina Marcela MenesesAún no hay calificaciones
- Revista Peruana de Medicina Experimental y Salud Pública 1726-4642Documento8 páginasRevista Peruana de Medicina Experimental y Salud Pública 1726-4642Rociio Del Carmen GonzalezAún no hay calificaciones
- Rejilla Embarazo EctópicoDocumento9 páginasRejilla Embarazo EctópicoJulissa GonzalesAún no hay calificaciones
- Remedios Naturales para Quistes en Los OvariosDocumento8 páginasRemedios Naturales para Quistes en Los OvariosPatty PoloAún no hay calificaciones
- 5 Embarazo EctopicoDocumento12 páginas5 Embarazo EctopicoLiz Torres ArteagaAún no hay calificaciones
- La MenstruaciónDocumento6 páginasLa MenstruaciónUhinnay YuAún no hay calificaciones
- Genitales Externos FemeninosDocumento17 páginasGenitales Externos FemeninosMariana Ramos GarciaAún no hay calificaciones
- Aspectos Fisiologicos de La Lactancia MaternaDocumento2 páginasAspectos Fisiologicos de La Lactancia Maternaluliarjol3Aún no hay calificaciones