También podría gustarte
- Origen Embriológico de RiñonDocumento1 páginaOrigen Embriológico de RiñonGlenn GarcíaAún no hay calificaciones
- Tercera A Octava Semana Del Desarrollo EmbrionarioDocumento124 páginasTercera A Octava Semana Del Desarrollo EmbrionarioVictor Guevara100% (7)
- Guia 14-15Documento10 páginasGuia 14-15adriana toainga vallin100% (1)
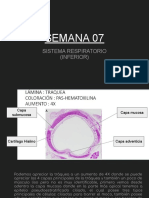
- Histología - Semana 07Documento25 páginasHistología - Semana 07Sharon GuevaraAún no hay calificaciones
- Embriologia Del Sistema DigestivoDocumento45 páginasEmbriologia Del Sistema DigestivoJcaminoAún no hay calificaciones
- Histología RiñónDocumento7 páginasHistología RiñónSneider Alexander Garcia TascoAún no hay calificaciones
- CorazónDocumento10 páginasCorazónJL Díaz0% (1)
- Embriologia Del Sistema CardiovascularDocumento32 páginasEmbriologia Del Sistema CardiovascularJcaminoAún no hay calificaciones
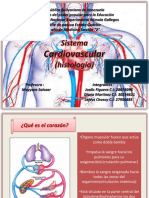
- Sistema Cardiovascular HistologiaDocumento26 páginasSistema Cardiovascular HistologiaAlbany ArteagaAún no hay calificaciones
- 0 - Bilirrubina, Grupo 3Documento52 páginas0 - Bilirrubina, Grupo 3Andrea Parodi VegaAún no hay calificaciones
- Biofisica de La Respiracion PDFDocumento68 páginasBiofisica de La Respiracion PDFClaudia Ovasco Rivera100% (1)
- Organoides CelularesDocumento8 páginasOrganoides CelularesSilvana StarAún no hay calificaciones
- Tipos de VenasDocumento14 páginasTipos de VenasIrene Sarai Horta ChiAún no hay calificaciones
- Laboratorio de TrematodosDocumento2 páginasLaboratorio de TrematodosLeslie VegaAún no hay calificaciones
- 2da Semana de DesarrolloDocumento6 páginas2da Semana de Desarrolloraul escobarAún no hay calificaciones
- Desarrollo de Vellosidades CorionicasDocumento11 páginasDesarrollo de Vellosidades CorionicasAlexia AlvaradoAún no hay calificaciones
- Histología Del Aparato Respiratorio InferiorDocumento24 páginasHistología Del Aparato Respiratorio InferiorJeanett Velásquez CorralesAún no hay calificaciones
- Resumen de La TraqueaDocumento12 páginasResumen de La TraqueaAlexis Andrade IbarraAún no hay calificaciones
- Cavidades CorporalesDocumento3 páginasCavidades CorporalesHeidi RamosAún no hay calificaciones
- Embriología y Anatomía Del Aparato Reproductor FemeninoDocumento17 páginasEmbriología y Anatomía Del Aparato Reproductor FemeninoCeci De La Flor CatacoraAún no hay calificaciones
- 3ro Aparato CirculatorioDocumento2 páginas3ro Aparato CirculatorioRuben PumaAún no hay calificaciones
- Protozoos CiliadosDocumento10 páginasProtozoos CiliadosKings Louis GeorgeAún no hay calificaciones
- Histologia de La PlacentaDocumento6 páginasHistologia de La PlacentaRepuesto BuenoAún no hay calificaciones
- La Formación de Mórula, Blastocisto y GastrulaciónDocumento21 páginasLa Formación de Mórula, Blastocisto y GastrulaciónMarlene ToledanoAún no hay calificaciones
- Genitales ExternosDocumento7 páginasGenitales ExternosLuisCarlosPerezAlvaradoAún no hay calificaciones
- Actividad 11 TallerSeñalizacionCelular-InteracciónCélulaCélulaDocumento9 páginasActividad 11 TallerSeñalizacionCelular-InteracciónCélulaCélulaAndrés FelipeAún no hay calificaciones
- Primera Semana de Desarrollo EmbrionarioDocumento6 páginasPrimera Semana de Desarrollo EmbrionarioMishel LopezAún no hay calificaciones
- Membrana CelularDocumento49 páginasMembrana CelularManuel Enrique Arana TrujilloAún no hay calificaciones
- EpidídimoDocumento5 páginasEpidídimoAnnaAún no hay calificaciones
- Vii CuestionarioDocumento4 páginasVii CuestionarioGabriela Mamani TrujilloAún no hay calificaciones
- Funciones Reproductoras y Hormonales Masculinas (Y Función de La Glándula PinealDocumento78 páginasFunciones Reproductoras y Hormonales Masculinas (Y Función de La Glándula PinealJenny AndreinaAún no hay calificaciones
- Cavidad Corporal, Mesentericos y DiafragmaDocumento12 páginasCavidad Corporal, Mesentericos y DiafragmaenriqueAún no hay calificaciones
- Las Tres Capas EmbrionariasDocumento28 páginasLas Tres Capas EmbrionariasWilliam Mullins100% (1)
- Compuestos NitrogenadosDocumento192 páginasCompuestos NitrogenadosMilton IsraelAún no hay calificaciones
- Aparato Genital Masculino MañanaDocumento9 páginasAparato Genital Masculino MañanaHuamani Flores JhAún no hay calificaciones
- Laringe Y TRAQUEADocumento22 páginasLaringe Y TRAQUEAAlejandra GonzalesAún no hay calificaciones
- NEOPLASIADocumento28 páginasNEOPLASIAGuiselle Andrea Verástegui BaldárragoAún no hay calificaciones
- Estructura y Funcion Del Nucleo CelularDocumento13 páginasEstructura y Funcion Del Nucleo CelularJohn Edgard Tuni HuallpaAún no hay calificaciones
- Unidad Ii. Cadena Respiratoria y Fosforilacion Oxidativa PDFDocumento34 páginasUnidad Ii. Cadena Respiratoria y Fosforilacion Oxidativa PDFMafer AraujoAún no hay calificaciones
- Mapas ConceptualesDocumento5 páginasMapas ConceptualesJerson BenturaAún no hay calificaciones
- AldosteronaDocumento19 páginasAldosteronakpumskwAún no hay calificaciones
- Capítulo 82 ExpoDocumento55 páginasCapítulo 82 Expoanon_216727310% (1)
- Piel FuncionesDocumento4 páginasPiel FuncionesMarissa PérezAún no hay calificaciones
- Órganos HematopoyéticosDocumento17 páginasÓrganos HematopoyéticosKaren OlveraAún no hay calificaciones
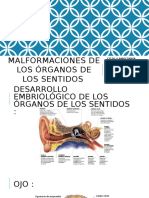
- Malformaciones de Los Órganos de Los SentidosDocumento29 páginasMalformaciones de Los Órganos de Los SentidosMilagros Saldaña CumpaAún no hay calificaciones
- Embriología Del Aparato Reproductor FemeninoDocumento9 páginasEmbriología Del Aparato Reproductor FemeninoAlberty RodriguezAún no hay calificaciones
- Embriogenesis de La Cuarta Semana A La Octava SemanaDocumento55 páginasEmbriogenesis de La Cuarta Semana A La Octava SemanaZOfii SnhzAún no hay calificaciones
- Histología de Los Ganglios LinfáticosDocumento10 páginasHistología de Los Ganglios LinfáticosDiana Marcela AcostaAún no hay calificaciones
- Capitulo 7 - Langman - Cavidades CorporalesDocumento3 páginasCapitulo 7 - Langman - Cavidades Corporalesmaria jose sanchez gilAún no hay calificaciones
- Mecanismos Basicos de La Funcion RenalDocumento3 páginasMecanismos Basicos de La Funcion RenalJuan Jose BarretoAún no hay calificaciones
- Respiratorio Bronquios y BronquiolosDocumento15 páginasRespiratorio Bronquios y BronquiolosLily MedinaAún no hay calificaciones
- El Tubo Intestinal y Las Cavidades CorporalesDocumento48 páginasEl Tubo Intestinal y Las Cavidades CorporalesUlises García Arellano100% (7)
- MFH Iii - Ao 11Documento45 páginasMFH Iii - Ao 11CAngeles S Subero100% (1)
- Sistema Cardiovascular. EMBRIOLOGIA PDFDocumento10 páginasSistema Cardiovascular. EMBRIOLOGIA PDFDavid De La RosaAún no hay calificaciones
- Tejido LinfoideDocumento4 páginasTejido Linfoided0ugy100% (2)
- Primera Semana de GestaciónDocumento3 páginasPrimera Semana de GestaciónAmal CalcagniAún no hay calificaciones
- Arterias ElásticasDocumento5 páginasArterias ElásticasKaren Geomara Naranjo LaraAún no hay calificaciones
- Circulacion FetalDocumento16 páginasCirculacion FetalNaddya BP100% (1)
- Embriologia: Segunda y Tercera Semana de DesarrolloDocumento20 páginasEmbriologia: Segunda y Tercera Semana de DesarrolloMedSchoolStuff100% (3)
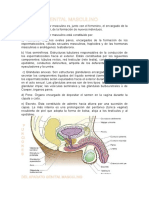
- Desarrollo Embrionario Del Aparato GenitourinarioDocumento23 páginasDesarrollo Embrionario Del Aparato Genitourinarionahideli sarahi ochoa barajasAún no hay calificaciones
- FUENTEOVEJUNADocumento21 páginasFUENTEOVEJUNAverolpz22gmailcom100% (1)
- Contrato en Materia LaboralDocumento3 páginasContrato en Materia Laboralmarcela badilloAún no hay calificaciones
- Evaluacion 3 IndividualDocumento5 páginasEvaluacion 3 IndividualJaviera IgnaciaAún no hay calificaciones
- Seder de Pesaj Vision CabalistaDocumento37 páginasSeder de Pesaj Vision CabalistaRoberto Molina100% (2)
- El Abrazo de OsoDocumento1 páginaEl Abrazo de OsoMaximiliano ImazAún no hay calificaciones
- 5 Trucos para Conseguir Un Estudio InteligenteDocumento29 páginas5 Trucos para Conseguir Un Estudio InteligenteArnolf Seminario ValladaresAún no hay calificaciones
- Literaturagauchesca 131218131300 Phpapp02 PDFDocumento9 páginasLiteraturagauchesca 131218131300 Phpapp02 PDFMelisa BenelliAún no hay calificaciones
- Tipos de Empresas en ColombiaDocumento2 páginasTipos de Empresas en ColombiaLuz JaraAún no hay calificaciones
- Educaion en Cuba Santos SoubalDocumento22 páginasEducaion en Cuba Santos SoubalAlberto Christin0% (1)
- Ramas CCPP FilosofíaDocumento3 páginasRamas CCPP FilosofíaJesús Esquerre NeyraAún no hay calificaciones
- Ficha de Actividades Proyecto 4 - MedioDocumento3 páginasFicha de Actividades Proyecto 4 - MedioAdri Baquez ChávezAún no hay calificaciones
- Diagnóstico 5toDocumento4 páginasDiagnóstico 5toMorena CanoAún no hay calificaciones
- Proyecto Final Gerencia Financiera 2Documento50 páginasProyecto Final Gerencia Financiera 2David NunezAún no hay calificaciones
- Escuela Nacional Superior Autonoma de Bellas Artes en JaujaDocumento22 páginasEscuela Nacional Superior Autonoma de Bellas Artes en JaujaEncuentro nacional de arte, diversidad cultural y educación (Perú)100% (2)
- Relacion Hispano Indigenas en ChileDocumento10 páginasRelacion Hispano Indigenas en ChileMelissa J. Moyano GuerraAún no hay calificaciones
- Casa Del Moral en ArequipaDocumento6 páginasCasa Del Moral en ArequipaYuli Gabriela Choque PeñalozaAún no hay calificaciones
- Los Fundamentos de La Administracion Del Trabajo PDFDocumento122 páginasLos Fundamentos de La Administracion Del Trabajo PDFCarlos Gavilanes TamayoAún no hay calificaciones
- Batch ProcessDocumento14 páginasBatch ProcessMargaret Lissette MorenoAún no hay calificaciones
- Diagnóstico Socioeducativo para La Planeación y Evaluación de La Gestión en Nuestra EscuelaDocumento1 páginaDiagnóstico Socioeducativo para La Planeación y Evaluación de La Gestión en Nuestra EscuelaMarcelino PoolAún no hay calificaciones
- Incendia Mi AlmaDocumento1 páginaIncendia Mi AlmaLili RiveraAún no hay calificaciones
- SESIÓN DE APRENDIZAJ #1 - 3 Y 4 Grado - I - EADocumento4 páginasSESIÓN DE APRENDIZAJ #1 - 3 Y 4 Grado - I - EAgaby sotoAún no hay calificaciones
- FiltrosDocumento10 páginasFiltrosJoel Jesus Perez UgarteAún no hay calificaciones
- Manual de Pediatria PDFDocumento518 páginasManual de Pediatria PDFRomina100% (1)
- Apuntes - Cap 3Documento10 páginasApuntes - Cap 3Lidia Ines Chavarria VasquezAún no hay calificaciones
- Plan de Emergencias de La Ips Sentir BienDocumento27 páginasPlan de Emergencias de La Ips Sentir BienEnriqueJaimeRomeroSolanoAún no hay calificaciones
- TAREA No 2Documento3 páginasTAREA No 2angela cruzAún no hay calificaciones
- Rivera La Revolución Es Un Sueño EternoDocumento19 páginasRivera La Revolución Es Un Sueño EternoAndrea PortugalAún no hay calificaciones
- Elasticidad Teo-ProblemasDocumento5 páginasElasticidad Teo-ProblemasFelipe SabelotoAún no hay calificaciones
- Caso de Exito ZARADocumento4 páginasCaso de Exito ZARAKatherin Quiñonez100% (1)
- PIMIENTADocumento1 páginaPIMIENTAManuel HarazemAún no hay calificaciones