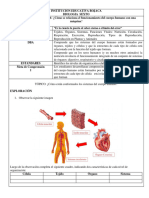
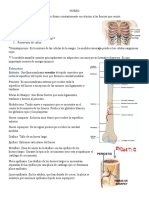
También podría gustarte
- Las células, Constructoras de vida: Cells: Constructing Living ThingsDe EverandLas células, Constructoras de vida: Cells: Constructing Living ThingsCalificación: 3.5 de 5 estrellas3.5/5 (2)
- Máquinas vivientes: ¿Cómo se mueven las células?De EverandMáquinas vivientes: ¿Cómo se mueven las células?Calificación: 5 de 5 estrellas5/5 (1)
- Lab 5-Histología AnimalDocumento11 páginasLab 5-Histología AnimalEdgar MartinezAún no hay calificaciones
- ORGANIZACIÓN Y FUNCION DE ORGANISMOS PLURICELULAR InvestigacionDocumento23 páginasORGANIZACIÓN Y FUNCION DE ORGANISMOS PLURICELULAR InvestigacionSulma HernandezAún no hay calificaciones
- A Conectivo Propio PDFDocumento13 páginasA Conectivo Propio PDFamarilis alejandraAún no hay calificaciones
- Ensayo LaboratorioDocumento16 páginasEnsayo LaboratorioAlpi NitoAún no hay calificaciones
- A Conectivo CartilagoDocumento17 páginasA Conectivo Cartilagoartenar2bernalmAún no hay calificaciones
- Tejidos TeoríaDocumento8 páginasTejidos TeoríaAaron Alexander Ceballos MorochoAún no hay calificaciones
- A Conectivo OseoDocumento16 páginasA Conectivo Oseoartenar2bernalmAún no hay calificaciones
- Tejido (Biología)Documento5 páginasTejido (Biología)nero ramoAún no hay calificaciones
- 1.-Tejidos BásicosDocumento11 páginas1.-Tejidos BásicosAlejandro B. SilvaAún no hay calificaciones
- A Conectivo SangreDocumento12 páginasA Conectivo Sangreartenar2bernalmAún no hay calificaciones
- Tejido VegetalDocumento3 páginasTejido VegetalMorales MoralesAún no hay calificaciones
- Documentos Johana 44Documento9 páginasDocumentos Johana 44Narvis BenavidedAún no hay calificaciones
- Caratula Colegio Edad de Oro TejidosDocumento10 páginasCaratula Colegio Edad de Oro TejidosJassel SantosAún no hay calificaciones
- Cedva 5Documento19 páginasCedva 5Andrea Jimena Sosa CanoAún no hay calificaciones
- Organización TisularDocumento8 páginasOrganización Tisularherminson pradaAún no hay calificaciones
- Laboratorio Biologia Tejidos AnimalesDocumento20 páginasLaboratorio Biologia Tejidos AnimalesVALENTINA CASTILLO GUERREROAún no hay calificaciones
- Organización FuncionalDocumento4 páginasOrganización FuncionalJasna ZovakAún no hay calificaciones
- HistologiaDocumento3 páginasHistologiaAriana Yusell Flores DiazAún no hay calificaciones
- Citología TRABAJO 1Documento11 páginasCitología TRABAJO 1Lau CubillasAún no hay calificaciones
- Reportedelaboratorio#3PA 114Documento17 páginasReportedelaboratorio#3PA 114Nahely PalaciosAún no hay calificaciones
- Trabajo de BiologíaDocumento6 páginasTrabajo de Biologíadl1956736Aún no hay calificaciones
- Celula Animal 10moDocumento13 páginasCelula Animal 10moIris Yolany Paz de MendozaAún no hay calificaciones
- Trabajo LexDocumento78 páginasTrabajo LexJuan Carlos RuizAún no hay calificaciones
- UD2 - Tema 2 TejidosDocumento24 páginasUD2 - Tema 2 TejidosyuliacgonzalezAún no hay calificaciones
- TejidosDocumento4 páginasTejidosEdison G.Aún no hay calificaciones
- Guía de TejidosDocumento6 páginasGuía de TejidosLuisa FernandaAún no hay calificaciones
- Borrador Atlas CelularDocumento67 páginasBorrador Atlas CelularGuallichico EmilyAún no hay calificaciones
- Anatomia Humana Tema 1Documento14 páginasAnatomia Humana Tema 1Maryann AlemánAún no hay calificaciones
- Afi1 U2 EaDocumento4 páginasAfi1 U2 EaGabriela MoralesAún no hay calificaciones
- HISTOLOGIADocumento12 páginasHISTOLOGIAJessica YepezAún no hay calificaciones
- CUADERNILLO BIOLOGIA UPLaBDocumento44 páginasCUADERNILLO BIOLOGIA UPLaBRoro WhooAún no hay calificaciones
- Tejido Animal.Documento10 páginasTejido Animal.KimAún no hay calificaciones
- TejidoDocumento4 páginasTejidoNatsu SalamanderAún no hay calificaciones
- A Conectivo SangreDocumento10 páginasA Conectivo SangreMara De La EspriellaAún no hay calificaciones
- AnatomofisiopatologiaDocumento229 páginasAnatomofisiopatologiashick_19Aún no hay calificaciones
- Tarea1 TejidosDocumento12 páginasTarea1 TejidosNegrete NegreteAún no hay calificaciones
- Informe Laboratorio Morfofisiologia-08!04!2014Documento28 páginasInforme Laboratorio Morfofisiologia-08!04!2014Carmen A TorresAún no hay calificaciones
- Los TejidosDocumento4 páginasLos TejidosSamuelSandovalVillarAún no hay calificaciones
- Guia 5 Tejidos AnimalesDocumento10 páginasGuia 5 Tejidos AnimalesHenrry Lois ReAún no hay calificaciones
- Clase Semana 1 - Fisiología - Naomi Guerra Mero PDFDocumento9 páginasClase Semana 1 - Fisiología - Naomi Guerra Mero PDFNaomi Guerra MeroAún no hay calificaciones
- Politecnico GrancolombianoDocumento6 páginasPolitecnico GrancolombianoNANI PEKIN100% (1)
- Informe de AnatomiaDocumento9 páginasInforme de AnatomiaNaileine BarcenasAún no hay calificaciones
- A Epitelio GlandularDocumento22 páginasA Epitelio GlandularJose Ignacio Lopez GarciaAún no hay calificaciones
- Introducción Al Cuerpo HumanoDocumento5 páginasIntroducción Al Cuerpo HumanoMore ForestelloAún no hay calificaciones
- AnatomiagggyyuDocumento10 páginasAnatomiagggyyularrietommohazz 28Aún no hay calificaciones
- Tejidos AnimalesDocumento9 páginasTejidos AnimalesPatricia CastroAún no hay calificaciones
- Informe de Laboratorio NoDocumento19 páginasInforme de Laboratorio NoLina SánchezAún no hay calificaciones
- Guia Cuerpo HumanoDocumento13 páginasGuia Cuerpo HumanokarlakaraligsAún no hay calificaciones
- Nivel Tisular de Organización - Tejidos AnimalesDocumento24 páginasNivel Tisular de Organización - Tejidos AnimalessoleymiAún no hay calificaciones
- Tema 2. Duodécimo. Los TejidosDocumento6 páginasTema 2. Duodécimo. Los TejidosKatihuska BatistaAún no hay calificaciones
- Bailón J. Clasificación Histológica. Tejidos Básicos o Primarios, Características Generales.Documento7 páginasBailón J. Clasificación Histológica. Tejidos Básicos o Primarios, Características Generales.Juan BailonAún no hay calificaciones
- HISTOLOGÍADocumento1 páginaHISTOLOGÍARoxana Ticona CayllanteAún no hay calificaciones
- Tejido Conectivo EspecializadoDocumento15 páginasTejido Conectivo EspecializadoLuis SingerAún no hay calificaciones
- Cómo Se Organizan Las Células en Los Organismos PluricelularesDocumento5 páginasCómo Se Organizan Las Células en Los Organismos PluricelularesAdam BoydAún no hay calificaciones
- Importancia de La HistologíaDocumento6 páginasImportancia de La HistologíaGladys Ibeth Mancera TapieroAún no hay calificaciones
- A Epitelio Glandular PDFDocumento17 páginasA Epitelio Glandular PDFNicolás RuizAún no hay calificaciones
- Capitulo 12-Araneae BajoDocumento24 páginasCapitulo 12-Araneae BajoDavid Vergara MorenoAún no hay calificaciones
- Alvear Et Al. - Práctica 2Documento16 páginasAlvear Et Al. - Práctica 2David Vergara MorenoAún no hay calificaciones
- Buelvas Et Al. - Proyecto CampoDocumento10 páginasBuelvas Et Al. - Proyecto CampoDavid Vergara MorenoAún no hay calificaciones
- Buelvas Et Al - Proyecto LabDocumento9 páginasBuelvas Et Al - Proyecto LabDavid Vergara MorenoAún no hay calificaciones
- PCR VirtualDocumento9 páginasPCR VirtualDavid Vergara MorenoAún no hay calificaciones
- Lab 1 - David VergaraDocumento4 páginasLab 1 - David VergaraDavid Vergara MorenoAún no hay calificaciones
- Extracción de ADN - Protocolo CaseroDocumento3 páginasExtracción de ADN - Protocolo CaseroDavid Vergara MorenoAún no hay calificaciones
- Paper Anatomia Del PeriodontoDocumento11 páginasPaper Anatomia Del PeriodontoLesly HerreraAún no hay calificaciones
- Discre ArtDocumento5 páginasDiscre ArtRonald Acarapi MuruchiAún no hay calificaciones
- Planos Tipicos de Corte para RevisarDocumento2 páginasPlanos Tipicos de Corte para RevisarPATRICK ALCIVARAún no hay calificaciones
- Resultado 202305101081Documento3 páginasResultado 202305101081Akemi TDAún no hay calificaciones
- Tejido Animal para Quinto Grado de PrimariaDocumento8 páginasTejido Animal para Quinto Grado de PrimariaEdgar FloresAún no hay calificaciones
- Manual de TincionesDocumento26 páginasManual de TincionesJorge Luis PS83% (6)
- Examen AnatomiaDocumento2 páginasExamen AnatomiaandreaAún no hay calificaciones
- CP Micro2Documento51 páginasCP Micro2Gerardo FloresAún no hay calificaciones
- Origen de La Tecnica Histologica en VenezuelaDocumento30 páginasOrigen de La Tecnica Histologica en Venezuelaclaudia BlandenierAún no hay calificaciones
- ENSAYO DE LA CELULA - BioquimicaDocumento4 páginasENSAYO DE LA CELULA - BioquimicaAlisson HernándezAún no hay calificaciones
- Clase#2 Dotación Cromosomica.Documento3 páginasClase#2 Dotación Cromosomica.Amy Dayana Cedeño ChávezAún no hay calificaciones
- Tejido Cartilajinoso Mapa ConceptualDocumento1 páginaTejido Cartilajinoso Mapa ConceptualAran VeraAún no hay calificaciones
- BiiologiaDocumento3 páginasBiiologiaClaudia Navarro JuarezAún no hay calificaciones
- Tejido NerviosoDocumento70 páginasTejido Nerviosoapi-3699557100% (6)
- Histo. Semana 3. Practica. Tejido ConectivoDocumento27 páginasHisto. Semana 3. Practica. Tejido ConectivoArieth G-aAún no hay calificaciones
- M1-C2 - Tejidos Básicos IDocumento30 páginasM1-C2 - Tejidos Básicos IEduardo JofreAún no hay calificaciones
- Actividad CelularDocumento20 páginasActividad CelularConstanza MaureiraAún no hay calificaciones
- HuesoDocumento7 páginasHuesogabrielAún no hay calificaciones
- Práctica 6. Tejidos VegetalesDocumento12 páginasPráctica 6. Tejidos VegetalesLaura Velásquez0% (1)
- Tipo de Orden Número ReqDocumento3 páginasTipo de Orden Número ReqEliamAún no hay calificaciones
- Manuel BetancourthDocumento2 páginasManuel BetancourthGabriel Evans AlvaradoAún no hay calificaciones
- CITOEQUELETODocumento3 páginasCITOEQUELETOEliam G. SánchezAún no hay calificaciones
- Tejido Muscular EUTM 2019Documento50 páginasTejido Muscular EUTM 2019sitiosdaniumtaAún no hay calificaciones
- FisiologíaDocumento5 páginasFisiologíaLucero Sonaly Mestanza LópezAún no hay calificaciones
- Estructura de La Celula Vegetal.Documento4 páginasEstructura de La Celula Vegetal.Renato GonzalezAún no hay calificaciones
- La Celula y Sus PartesDocumento2 páginasLa Celula y Sus Partessandy lAún no hay calificaciones
- Evaluación de La Calidad de CPQ Provenientes de Sangre Total en El Hemocentro de La Cruz Roja Ecuatoriana 2014Documento91 páginasEvaluación de La Calidad de CPQ Provenientes de Sangre Total en El Hemocentro de La Cruz Roja Ecuatoriana 2014williams luis arana florianoAún no hay calificaciones
- Cuestionario de Biología - Biomedicina 3er CicloDocumento8 páginasCuestionario de Biología - Biomedicina 3er CicloMacas AnyeloAún no hay calificaciones
- ERITROPOYESISDocumento19 páginasERITROPOYESISMaria PaulinaAún no hay calificaciones
- Domino de La CelulaDocumento1 páginaDomino de La Celulaingrid katherin rivas cabrales100% (1)