También podría gustarte
- Triptico CromosomasDocumento2 páginasTriptico CromosomasTibulo Vera67% (3)
- Fisiología Del Sistema EndocrinoDocumento96 páginasFisiología Del Sistema EndocrinoUSMP FN ARCHIVOSAún no hay calificaciones
- Receptores HormonalesDocumento29 páginasReceptores HormonalesTeresa MatusAún no hay calificaciones
- Regulacion Hormonal Resumen IMPRIMIRDocumento8 páginasRegulacion Hormonal Resumen IMPRIMIRTania CabreraAún no hay calificaciones
- Bioquímica - Módulo 2: Tema 1: Metabolismo de InformaciónDocumento89 páginasBioquímica - Módulo 2: Tema 1: Metabolismo de InformaciónAlejandra PaolaAún no hay calificaciones
- BIOSEÑALIZACIÓNDocumento11 páginasBIOSEÑALIZACIÓNelisa ponce de leonAún no hay calificaciones
- Mecanismos de Accion HormonalDocumento4 páginasMecanismos de Accion HormonalCarlitos SanchezAún no hay calificaciones
- Seminario de HormonasDocumento17 páginasSeminario de HormonasAnonymous eKu3ujsvQg75% (4)
- Fisiologia Endocrina EditDocumento7 páginasFisiologia Endocrina EditPia CortesAún no hay calificaciones
- SEÑALIZACIONDocumento16 páginasSEÑALIZACIONHaritchet 15Aún no hay calificaciones
- Exposicion Fisiologia.Documento24 páginasExposicion Fisiologia.patricia cubileteAún no hay calificaciones
- 10 Transducción de Señales PDFDocumento32 páginas10 Transducción de Señales PDFIrene CarballoAún no hay calificaciones
- Discusion 4 CompletaDocumento26 páginasDiscusion 4 CompletaJorge HernándezAún no hay calificaciones
- Fisiologia Sistema e 238504 Downloadable 1971837Documento11 páginasFisiologia Sistema e 238504 Downloadable 1971837elizabethtoribio22Aún no hay calificaciones
- Sistema Endocrino 137681 Downloable 910132Documento8 páginasSistema Endocrino 137681 Downloable 910132López Espinoza Livia FernandaAún no hay calificaciones
- Resumen HORMONASDocumento8 páginasResumen HORMONASmartinamorelvulliez12345Aún no hay calificaciones
- Tema 47Documento6 páginasTema 47gabrielmd858Aún no hay calificaciones
- Accion Hormonal y Transducción de La SeñalDocumento5 páginasAccion Hormonal y Transducción de La SeñalRuth Elizabeth Ascona VeraAún no hay calificaciones
- Bioquimica - Semana 5Documento9 páginasBioquimica - Semana 5Miguel Milano FloresAún no hay calificaciones
- Sistema EndocrinoDocumento3 páginasSistema Endocrinomobufe2Aún no hay calificaciones
- Fisiologia Sistema e 238504 Downloadable 1654242Documento11 páginasFisiologia Sistema e 238504 Downloadable 1654242DANNA PAOLA LEYVA CANTUAún no hay calificaciones
- Endocrino GeneralDocumento31 páginasEndocrino GeneralJose CabreraAún no hay calificaciones
- ENDOCRINO, Anexo Final A Cap Del GuytonDocumento14 páginasENDOCRINO, Anexo Final A Cap Del GuytoncristinaaduartefacultadAún no hay calificaciones
- Señalización IntracelularDocumento3 páginasSeñalización IntracelularALONDRA MONTSERRAT GRADILLA SILVAAún no hay calificaciones
- 2) Introduccion A La EndocrinologiaDocumento47 páginas2) Introduccion A La EndocrinologiaMarlon RangelAún no hay calificaciones
- Blancos Farmacológicos en La Membrana CelularDocumento3 páginasBlancos Farmacológicos en La Membrana CelularLuis López Medina.Aún no hay calificaciones
- Vitaminas Quimica 2Documento6 páginasVitaminas Quimica 2Javier PerezAún no hay calificaciones
- Clase 4 Tipos de ReceptoresDocumento13 páginasClase 4 Tipos de ReceptoresSonicAún no hay calificaciones
- Receptores Acoplados A Proteina GDocumento3 páginasReceptores Acoplados A Proteina GRossana RosadoAún no hay calificaciones
- Capitulo 74 y 75Documento90 páginasCapitulo 74 y 75Bruno SoaresAún no hay calificaciones
- 2) FarmacodinamiaDocumento13 páginas2) FarmacodinamiaMilagros SilvaAún no hay calificaciones
- BioseñalizaciónDocumento64 páginasBioseñalizaciónEdson EstelaAún no hay calificaciones
- Fisiologia Semana 4Documento11 páginasFisiologia Semana 4nicollgarsomAún no hay calificaciones
- HormonaDocumento8 páginasHormonaOntiveros AylinAún no hay calificaciones
- EXPOSICIONDocumento42 páginasEXPOSICIONJade MendozaAún no hay calificaciones
- Bioca CDI - Semana 1 HormonasDocumento9 páginasBioca CDI - Semana 1 HormonasOscar Abraham Aguilar PopocaAún no hay calificaciones
- Hormonas MaDocumento14 páginasHormonas MaAnahi NavarroAún no hay calificaciones
- FarmacodinamiaDocumento28 páginasFarmacodinamiaDanna GutiérrezAún no hay calificaciones
- HormonasDocumento33 páginasHormonasEsthefanny Lizeth Carrasco GarcésAún no hay calificaciones
- Hormonas y Sus Mecanismos de AcciónDocumento10 páginasHormonas y Sus Mecanismos de AcciónMariaAún no hay calificaciones
- Presentacion Medicina y Salud Ilustrativo Infantil CelesteDocumento38 páginasPresentacion Medicina y Salud Ilustrativo Infantil Celeste2022321072Aún no hay calificaciones
- Tema 5Documento6 páginasTema 5Erika Luna JiménezAún no hay calificaciones
- Hormonas: Dra. Fabiola Mendoza de Rodas Marzo 2023Documento52 páginasHormonas: Dra. Fabiola Mendoza de Rodas Marzo 2023Paolafer RivasAún no hay calificaciones
- Tema de HORMONASDocumento7 páginasTema de HORMONASAndrea DiazAún no hay calificaciones
- Repaso 2do Parcial ReproDocumento60 páginasRepaso 2do Parcial Reprojoandaly quirozAún no hay calificaciones
- Sistema EndocrinoDocumento17 páginasSistema EndocrinoVentas TupperwareAún no hay calificaciones
- Sistema EndocrinoDocumento10 páginasSistema EndocrinoDaniela Fernanda Ochoa SanchezAún no hay calificaciones
- Receptores en FarmacodinamiaDocumento18 páginasReceptores en FarmacodinamiaLucas VélezAún no hay calificaciones
- Fisiología Del Sistema EndocrinoDocumento95 páginasFisiología Del Sistema EndocrinoAllison GregoryAún no hay calificaciones
- Proteinas GDocumento16 páginasProteinas GAna Karina Tarco PerezAún no hay calificaciones
- 1 - Generalidades EndócrinoDocumento3 páginas1 - Generalidades EndócrinoCeleste GuanaisAún no hay calificaciones
- Tarea 6 de Anatomia y Fisiologia Del Sistema NerviosoDocumento28 páginasTarea 6 de Anatomia y Fisiologia Del Sistema NerviosoMASSIEL AMADOR GREEN0% (1)
- EndocrinologiaDocumento30 páginasEndocrinologiaMorejon Aguila Jose AlejandroAún no hay calificaciones
- Accion de Los Agentes Neurohumorales Sobre La Membrana y Sobre Los Organos Intracitoplasmaticos Avalos Servin Wendolyn MonserratDocumento4 páginasAccion de Los Agentes Neurohumorales Sobre La Membrana y Sobre Los Organos Intracitoplasmaticos Avalos Servin Wendolyn MonserratVictorJMeloAún no hay calificaciones
- Mecanismos de Comunicación Celular y Regulacion...Documento28 páginasMecanismos de Comunicación Celular y Regulacion...Keniia De La VegaAún no hay calificaciones
- Sistema Endrocino GeneralidadesDocumento4 páginasSistema Endrocino GeneralidadesMelissa MorenoAún no hay calificaciones
- Señalizacion HormonalDocumento8 páginasSeñalizacion HormonalGabriela Trocel MendezAún no hay calificaciones
- Discusión 4 CompletaDocumento30 páginasDiscusión 4 CompletaRobin SmithAún no hay calificaciones
- Endocrino Seminario 1Documento5 páginasEndocrino Seminario 1borellaazulAún no hay calificaciones
- Clase 5 - Bii - 2023Documento30 páginasClase 5 - Bii - 2023Dany MirellaAún no hay calificaciones
- 4B Examen I Bioestadística 2022Documento3 páginas4B Examen I Bioestadística 2022Aitana PiñaAún no hay calificaciones
- Mapa Glaucoma-Piña Delgado AitanaDocumento1 páginaMapa Glaucoma-Piña Delgado AitanaAitana PiñaAún no hay calificaciones
- Primer Parcial Medicina LaboralDocumento7 páginasPrimer Parcial Medicina LaboralAitana PiñaAún no hay calificaciones
- Caso Clínico 11 2.0Documento1 páginaCaso Clínico 11 2.0Aitana PiñaAún no hay calificaciones
- Mapa Conceptual Riesgos de Trabajo-Piña Delgado AitanaDocumento1 páginaMapa Conceptual Riesgos de Trabajo-Piña Delgado AitanaAitana PiñaAún no hay calificaciones
- Cartel-Analgesia Inmersion ColaDocumento1 páginaCartel-Analgesia Inmersion ColaAitana PiñaAún no hay calificaciones
- Mapa Retinopatia Diabetica-Piña Delgado AitanaDocumento1 páginaMapa Retinopatia Diabetica-Piña Delgado AitanaAitana PiñaAún no hay calificaciones
- Cuestionario 1-Piña Delgado AitanaDocumento2 páginasCuestionario 1-Piña Delgado AitanaAitana PiñaAún no hay calificaciones
- Resumen Articulo Nuevos Tratamientos para Gramnegativos-UAEM 5B-Pichardo, Pineda, PiñaDocumento2 páginasResumen Articulo Nuevos Tratamientos para Gramnegativos-UAEM 5B-Pichardo, Pineda, PiñaAitana PiñaAún no hay calificaciones
- Practica 15 Funcion PulmonarDocumento9 páginasPractica 15 Funcion PulmonarAitana PiñaAún no hay calificaciones
- Practica 17 TermoregulacionDocumento9 páginasPractica 17 TermoregulacionAitana PiñaAún no hay calificaciones
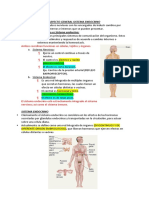
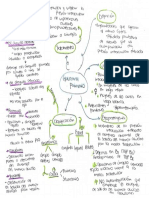
- Piña Aitana-Introducción A La Fisiología EndocrinaDocumento2 páginasPiña Aitana-Introducción A La Fisiología EndocrinaAitana PiñaAún no hay calificaciones
- Practica 14 Funcion RenalDocumento16 páginasPractica 14 Funcion RenalAitana PiñaAún no hay calificaciones
- Piña - Aitana - Actividad - 8. Tema - Ciclo Vital y Salud MentalDocumento25 páginasPiña - Aitana - Actividad - 8. Tema - Ciclo Vital y Salud MentalAitana PiñaAún no hay calificaciones
- Piña - Aitana - Actividad Practica Clínica 3. Episodio 2 "La Barrera". Hospital Lenox Hill de Nueva York EE - UU. Documental Sobre Medicina.Documento4 páginasPiña - Aitana - Actividad Practica Clínica 3. Episodio 2 "La Barrera". Hospital Lenox Hill de Nueva York EE - UU. Documental Sobre Medicina.Aitana PiñaAún no hay calificaciones
- Bioetica-Primera ClaseDocumento27 páginasBioetica-Primera ClaseAitana PiñaAún no hay calificaciones
- Caso Clinico Embarazo de 42 SDGDocumento2 páginasCaso Clinico Embarazo de 42 SDGAitana PiñaAún no hay calificaciones
- Division CelularDocumento8 páginasDivision CelularPia MedicinaAún no hay calificaciones
- Trabajo Práctico N°2 NutricionDocumento2 páginasTrabajo Práctico N°2 NutricionSergio BalmacedaAún no hay calificaciones
- Beta Oxidacion de Los Acid Grasos InsatuDocumento9 páginasBeta Oxidacion de Los Acid Grasos InsatuKatherin BeraunAún no hay calificaciones
- Secuencia Del Genoma de Yersinia Pestis KIM ResumenDocumento3 páginasSecuencia Del Genoma de Yersinia Pestis KIM ResumenvanessaAún no hay calificaciones
- Trabajo 1Documento8 páginasTrabajo 1Vanessa DuqueAún no hay calificaciones
- Degrabada Inmuno 05Documento25 páginasDegrabada Inmuno 05Enrique Moncada MapelliAún no hay calificaciones
- ¿Qué Es La Ingeniería Genética?: Cuaderno #4, Edición 2021Documento7 páginas¿Qué Es La Ingeniería Genética?: Cuaderno #4, Edición 2021Mildred MunguíaAún no hay calificaciones
- Wuolah Free TEMARIO BIOLOGIA CELULAR E INMUNOLOGIADocumento149 páginasWuolah Free TEMARIO BIOLOGIA CELULAR E INMUNOLOGIASergioAún no hay calificaciones
- Manual de CosDocumento248 páginasManual de CosfcalarconAún no hay calificaciones
- Escrito Relacion Video y OVAS1 y OVAS 2Documento2 páginasEscrito Relacion Video y OVAS1 y OVAS 2Yessica MorenoAún no hay calificaciones
- QPCRDocumento34 páginasQPCRLidiaAlejandraRiveraPonceAún no hay calificaciones
- La Fisión BinariaDocumento3 páginasLa Fisión BinariaRobert Mayta100% (1)
- Tipos de Arn RibosomalDocumento2 páginasTipos de Arn RibosomalMarcelitito VayitassAún no hay calificaciones
- Digitacion HematologiaDocumento4 páginasDigitacion HematologiaRodriguez J ElizabethAún no hay calificaciones
- PRS BQ 06 EnzimasDocumento40 páginasPRS BQ 06 Enzimasmario antonio b lazariniAún no hay calificaciones
- FundamentosDocumento36 páginasFundamentosDannia DelgadoAún no hay calificaciones
- Cien 11ºDocumento44 páginasCien 11ºMelissa D TorresAún no hay calificaciones
- Exposicion Adhesion CelularDocumento19 páginasExposicion Adhesion CelularKristel SolisAún no hay calificaciones
- Crucigrama CelularDocumento2 páginasCrucigrama CelularJesús Salvador Cervantes HigaredaAún no hay calificaciones
- Practica de Laboratorio Semana 2Documento11 páginasPractica de Laboratorio Semana 2Jhon GuerraAún no hay calificaciones
- ChaperonasDocumento7 páginasChaperonasOscar Cardona PantojaAún no hay calificaciones
- 4 Barreras y PRR - SazgDocumento33 páginas4 Barreras y PRR - SazgFernanda CanalesAún no hay calificaciones
- Clase 4 - Regulación Actividad EnzimáticaDocumento7 páginasClase 4 - Regulación Actividad EnzimáticaanaAún no hay calificaciones
- El Núcleo CelularDocumento6 páginasEl Núcleo CelularGiuliana GonzalezAún no hay calificaciones
- Citología ESMDocumento28 páginasCitología ESMELENA SANCHEZ MAURIAún no hay calificaciones
- Control de Calidad en Biologia MolecularDocumento65 páginasControl de Calidad en Biologia Molecular05-TL-HU-ERICK ANTONY BARZOLA GALDOSAún no hay calificaciones
- Informe de Laboratorio Italo GrottiniDocumento2 páginasInforme de Laboratorio Italo GrottiniBenjamin Alejandro Sanchez OlivazAún no hay calificaciones
- Cuestionario LecheDocumento5 páginasCuestionario Lechesandra Aguilera100% (1)
- Act BiomoleculasDocumento4 páginasAct BiomoleculasRoberto García OjedaAún no hay calificaciones