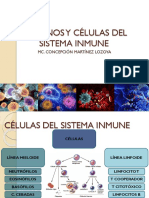
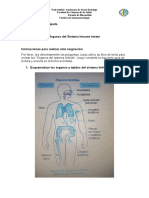
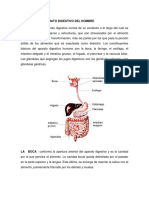
También podría gustarte
- Monografia (Completa)Documento18 páginasMonografia (Completa)Graciela IrmaAún no hay calificaciones
- Organos Linfoides Primarios y SecundariosDocumento3 páginasOrganos Linfoides Primarios y SecundariosKiimPossible100% (4)
- Órganos y Células Del Sistema InmuneDocumento50 páginasÓrganos y Células Del Sistema InmuneAngie ManningsAún no hay calificaciones
- 04 Inmunología Del Tracto DigestivoDocumento81 páginas04 Inmunología Del Tracto DigestivoSebastian Requena PasacheAún no hay calificaciones
- Tema Nº2 Inmunología ParasitariaDocumento55 páginasTema Nº2 Inmunología ParasitariaBrian CalleAún no hay calificaciones
- INGESTIÓNDocumento5 páginasINGESTIÓNminipabloxd1Aún no hay calificaciones
- Inmunidad Del Sistema DigestivoDocumento3 páginasInmunidad Del Sistema Digestivojuliana100% (5)
- 1 - Inmunidad y Mucosas 2016Documento49 páginas1 - Inmunidad y Mucosas 2016Daii YJonaAún no hay calificaciones
- Inmunidad en Las MucosasDocumento8 páginasInmunidad en Las MucosasSofia DreispielAún no hay calificaciones
- Aparato DigestivoDocumento4 páginasAparato DigestivoTania Tzab50% (2)
- Hormonas y Enzimas Digestivas EnvioDocumento2 páginasHormonas y Enzimas Digestivas EnvioKarlheinz Fernandez Bermudez100% (2)
- Tejido Linfoide Asociado A La MucosaDocumento3 páginasTejido Linfoide Asociado A La MucosaMaria Camila GonzálezAún no hay calificaciones
- Seminario InmunoDocumento8 páginasSeminario InmunoMar Vidal LópezAún no hay calificaciones
- Tarea 2.3 - Organos Del Sistema InmuneDocumento9 páginasTarea 2.3 - Organos Del Sistema InmuneRosmery Sánchez TejadaAún no hay calificaciones
- GastrofisiologiaDocumento14 páginasGastrofisiologiaMelanie EspinozaAún no hay calificaciones
- El Intestino Pieza Clave en El Sistema InmuneDocumento13 páginasEl Intestino Pieza Clave en El Sistema Inmuneluz garciaAún no hay calificaciones
- Competencia 4Documento6 páginasCompetencia 4jaircaleb12Aún no hay calificaciones
- Monografia GALTDocumento5 páginasMonografia GALTOmar Santiago RamirezAún no hay calificaciones
- Alvaro AvesDocumento25 páginasAlvaro AvesALVARO JOSE VILLAMIZAR JIMENEZAún no hay calificaciones
- Inmunologia Unidad 3Documento25 páginasInmunologia Unidad 3Cristhian DiazAún no hay calificaciones
- Sistema InmunitarioDocumento17 páginasSistema InmunitarioCarlos Zanns CarpioAún no hay calificaciones
- Sistema Inmune MucosasDocumento7 páginasSistema Inmune Mucosasabelhenarejos-1Aún no hay calificaciones
- Tema 6.1 Inmunidad de Las Mucosas.Documento47 páginasTema 6.1 Inmunidad de Las Mucosas.Nancy Liliana Bolivar GuizarAún no hay calificaciones
- Reporte de Lectura 8Documento3 páginasReporte de Lectura 8SherlynAún no hay calificaciones
- Respuesta Del Huésped A La InfecciónDocumento1 páginaRespuesta Del Huésped A La InfecciónCristian Emanuel Argueta ChicasAún no hay calificaciones
- 2-Sistema-Inmune-celular CompletoDocumento6 páginas2-Sistema-Inmune-celular CompletoGuille OliveraAún no hay calificaciones
- Inmunidad HumoralDocumento16 páginasInmunidad HumoralOscar Miguel Hernandez CastilloAún no hay calificaciones
- Organos Linfoides Del CerdoDocumento2 páginasOrganos Linfoides Del CerdoGustavo SteinbregerAún no hay calificaciones
- GALTDocumento1 páginaGALTaemoran115Aún no hay calificaciones
- OrganossistemainmuneDocumento16 páginasOrganossistemainmuneoktaviusmasterAún no hay calificaciones
- Taller InmunologiaDocumento15 páginasTaller Inmunologiaanon_230054309Aún no hay calificaciones
- Primera FaseDocumento35 páginasPrimera FaseKarla ApazaAún no hay calificaciones
- Micro Tarea 2Documento11 páginasMicro Tarea 2Jocelyn HdezAún no hay calificaciones
- MALTDocumento11 páginasMALTcarmen pitreAún no hay calificaciones
- Inmunidad Innata en El Tubo DigestivoDocumento5 páginasInmunidad Innata en El Tubo DigestivoJamil Litardo VillacresesAún no hay calificaciones
- INMUNAIRDocumento22 páginasINMUNAIRjob88264629100% (1)
- Inmunidad Especializada en Las Barreras Epiteliales y Tej PrivilegiadosDocumento33 páginasInmunidad Especializada en Las Barreras Epiteliales y Tej PrivilegiadosJenn Abad (I am a student)Aún no hay calificaciones
- Tejido Linfático Difuso y Nódulos LinfáticosDocumento2 páginasTejido Linfático Difuso y Nódulos LinfáticosJose Ortiz VermilionAún no hay calificaciones
- Células Del Sistema InmuneDocumento7 páginasCélulas Del Sistema InmuneSantiago HerreraAún no hay calificaciones
- Organos LinfoidesDocumento3 páginasOrganos LinfoidesEduardo FalcónAún no hay calificaciones
- Resumen de Inmunologia - Celulas y Organos Del Sistema InmuneDocumento7 páginasResumen de Inmunologia - Celulas y Organos Del Sistema InmuneEsmeralda HernándezAún no hay calificaciones
- Sistema Inmunologico (Geral 001)Documento60 páginasSistema Inmunologico (Geral 001)titoaymaraAún no hay calificaciones
- Trabajo MALTDocumento2 páginasTrabajo MALTYuki Chari Moco Saray50% (4)
- Tema 6 Inmuno 1era ParteDocumento14 páginasTema 6 Inmuno 1era ParteMaría Alvarado PachasAún no hay calificaciones
- Inmunidad de MucosasDocumento13 páginasInmunidad de MucosasIGNACIO SALVADOR FERNANDEZ GIRONAún no hay calificaciones
- Histofisiologia Del Timo-Sistema Rio LinfaticoDocumento10 páginasHistofisiologia Del Timo-Sistema Rio LinfaticoMicaela SecadaAún no hay calificaciones
- Adaptaciones ParasitariasDocumento4 páginasAdaptaciones ParasitariascriznahpAún no hay calificaciones
- Sistema InmuneDocumento3 páginasSistema InmuneZabrinaletAún no hay calificaciones
- Organos y Tejidos Implicados en La Respuesta InmuneDocumento8 páginasOrganos y Tejidos Implicados en La Respuesta Inmunelezc239Aún no hay calificaciones
- Caracteristicas Generales Del Sistema InmunologicoDocumento5 páginasCaracteristicas Generales Del Sistema InmunologicoWilfer Hilaquita Condori0% (2)
- Inmuno GastroDocumento69 páginasInmuno GastrodrAún no hay calificaciones
- Guia InmunologíaDocumento4 páginasGuia InmunologíaOriana MezaAún no hay calificaciones
- Inmunidad Intestinal.Documento77 páginasInmunidad Intestinal.fiorellaAún no hay calificaciones
- 06 Procesamiento y Presentacion Del Ag A Los Linfocitos T LecturaDocumento18 páginas06 Procesamiento y Presentacion Del Ag A Los Linfocitos T LecturaVanessa Tapia QuirozAún no hay calificaciones
- Inmunidad A Mucosas Parte 1Documento74 páginasInmunidad A Mucosas Parte 1Vanessa YapoAún no hay calificaciones
- Inmunologia 2.0Documento15 páginasInmunologia 2.0Elianny BriceñoAún no hay calificaciones
- Inmunidad Innata Primera Expo Sic IonDocumento9 páginasInmunidad Innata Primera Expo Sic IonLiz Xenia TerrazasAún no hay calificaciones
- Organos Linfoides Primarios y SegundariosDocumento12 páginasOrganos Linfoides Primarios y SegundariosKaira Zonard100% (2)
- Linfocitos T y B 2Documento28 páginasLinfocitos T y B 2Deyvis Andres Nieves FloresAún no hay calificaciones
- 06 Inmunidad de MucosasDocumento37 páginas06 Inmunidad de Mucosassilvia sandra sulca arangoAún no hay calificaciones
- IFAPESsistemalinfoide 1234Documento11 páginasIFAPESsistemalinfoide 1234Antonio Belmonte LealAún no hay calificaciones
- Organos Del Sistema InmuneDocumento20 páginasOrganos Del Sistema InmuneMontserrat Abigail Ordóñez MéndezAún no hay calificaciones
- Histologia Del Intestino DelgadoDocumento2 páginasHistologia Del Intestino DelgadoIhanabAún no hay calificaciones
- Preguntas Fisiologia DegestivaDocumento5 páginasPreguntas Fisiologia DegestivaPercy Huacarpuma CasimiroAún no hay calificaciones
- Aparato DigestivoDocumento7 páginasAparato DigestivoivanAún no hay calificaciones
- Biologia Agosto 2023 ResueltoDocumento152 páginasBiologia Agosto 2023 ResueltoCristina BritoAún no hay calificaciones
- Hoja de Trabajo-Componentes Del Sistema DigestivoDocumento4 páginasHoja de Trabajo-Componentes Del Sistema DigestivoRuben Daniel LeonAún no hay calificaciones
- Sistema Digestivo y Sistema Respiratorio Geupo 4Documento74 páginasSistema Digestivo y Sistema Respiratorio Geupo 4Yessy CalderonAún no hay calificaciones
- ABDOMENDocumento111 páginasABDOMENMafeNarvaezAún no hay calificaciones
- Proceso de Digestión de Los Alimentos.Documento4 páginasProceso de Digestión de Los Alimentos.Samantha SanchezAún no hay calificaciones
- Final NutricionDocumento131 páginasFinal NutricionJair Orlando Rocha VillalpandoAún no hay calificaciones
- Parasitos Internos OvinosDocumento13 páginasParasitos Internos OvinosValeria FloresAún no hay calificaciones
- Cuestionario Morfo.Documento6 páginasCuestionario Morfo.Sarita AbigailAún no hay calificaciones
- Intestino DelgadoDocumento3 páginasIntestino DelgadoJuan DavidAún no hay calificaciones
- Expo GuiaDocumento51 páginasExpo GuiaJorge Luis Griego HerreraAún no hay calificaciones
- Material de Apoyo Asistencia A Procedimientos Medico QuirurgicosDocumento81 páginasMaterial de Apoyo Asistencia A Procedimientos Medico Quirurgicosgeraldine cardonaAún no hay calificaciones
- Digest IvoDocumento29 páginasDigest IvoMax Edwin Gorena EspinozaAún no hay calificaciones
- Amebiasis y GiardiasisDocumento10 páginasAmebiasis y GiardiasisVanesa Giraldo ZuluagaAún no hay calificaciones
- Aparato Digestivo Del HombreDocumento3 páginasAparato Digestivo Del HombrePipe HenaoAún no hay calificaciones
- Intestino DelgadoDocumento10 páginasIntestino DelgadoJavier ApazaAún no hay calificaciones
- Qué Pasa Por Dentro Cuando ComemosDocumento3 páginasQué Pasa Por Dentro Cuando ComemosJosé HdezAún no hay calificaciones
- Documentode Estudio Sistema DigestivoDocumento13 páginasDocumentode Estudio Sistema DigestivoLudy mariamAún no hay calificaciones
- Anatomia Del Sistema DigestivoDocumento63 páginasAnatomia Del Sistema DigestivoJosue Orozco0% (1)
- Descripción Morfológica de Cotylophoron Cotylophorum Colombia PDFDocumento11 páginasDescripción Morfológica de Cotylophoron Cotylophorum Colombia PDFHarold Christopher Cortez CercadoAún no hay calificaciones
- Sistema Dijestivo 2019Documento23 páginasSistema Dijestivo 2019Laura100% (1)
- Desarrollo Fisico de 0 - 6 AñosDocumento5 páginasDesarrollo Fisico de 0 - 6 Añosyasmin315Aún no hay calificaciones
- Aves TareaDocumento24 páginasAves TareaAlexia AraujoAún no hay calificaciones
- Estudio de La Malabsorción de Sales BiliaresDocumento1 páginaEstudio de La Malabsorción de Sales BiliaresNataly QuevedoAún no hay calificaciones
- Catalogo Base 2019 CDCDocumento73 páginasCatalogo Base 2019 CDCdrpegueroAún no hay calificaciones