También podría gustarte
- 4 Año Guia 1 Biologia Momento 3Documento8 páginas4 Año Guia 1 Biologia Momento 3Ramos RamonAún no hay calificaciones
- Porifera y PlacozoaDocumento21 páginasPorifera y PlacozoaDiego NapánAún no hay calificaciones
- Invertebrados: esponjas y artrópodosDocumento62 páginasInvertebrados: esponjas y artrópodosdjmauro25Aún no hay calificaciones
- Trabajo Anfibios y Reptiles.Documento32 páginasTrabajo Anfibios y Reptiles.Caroliina Morales80% (5)
- Parasitos en PecesDocumento6 páginasParasitos en PecesFabián Cortés LilloAún no hay calificaciones
- Hábitos alimenticios de moluscos poliplacóforosDocumento42 páginasHábitos alimenticios de moluscos poliplacóforosHomero AlvarezAún no hay calificaciones
- Fatouh - 2do Ano - Reino Protista - Apunte Teorico y Cuestionario 2009Documento15 páginasFatouh - 2do Ano - Reino Protista - Apunte Teorico y Cuestionario 2009api-3824772Aún no hay calificaciones
- Informa de La Practica3. Introduccion A Los Poriferos.Documento9 páginasInforma de La Practica3. Introduccion A Los Poriferos.Rodexy Esther Cuevas NovasAún no hay calificaciones
- Pia Final BinaDocumento22 páginasPia Final BinaLizbette RiveraAún no hay calificaciones
- Informe Documentado, Porífera y CnidariaDocumento8 páginasInforme Documentado, Porífera y CnidariaKeyla Michel GarciaAún no hay calificaciones
- Características de Platyhelminthes, Mollusca y ArthropodaDocumento14 páginasCaracterísticas de Platyhelminthes, Mollusca y ArthropodaDiego NapánAún no hay calificaciones
- Origen y evolución de los anfibios desde el DevónicoDocumento9 páginasOrigen y evolución de los anfibios desde el DevónicoXavier Valencia100% (1)
- Sinopsis Taxonómica de AnthropodaDocumento9 páginasSinopsis Taxonómica de AnthropodaAngie LizethAún no hay calificaciones
- Sesion Teorica 03Documento8 páginasSesion Teorica 03Daniela NuñezAún no hay calificaciones
- Bina Pia2Documento15 páginasBina Pia2Lizbette RiveraAún no hay calificaciones
- Informe 5 ZooDocumento13 páginasInforme 5 ZooDiego NapánAún no hay calificaciones
- Forma de Poriferos PDFDocumento10 páginasForma de Poriferos PDFJavier Polo NúñezAún no hay calificaciones
- ProtistaDocumento21 páginasProtistawaldrapa29Aún no hay calificaciones
- Acuario Mundo MarinoDocumento16 páginasAcuario Mundo MarinoSandra Milena Guerra HernándezAún no hay calificaciones
- Informe No. 1 ProtozoariosDocumento12 páginasInforme No. 1 ProtozoariosAngel Daniel Ausecha VivasAún no hay calificaciones
- Los ForaminiferosDocumento21 páginasLos Foraminiferosmochavez100% (1)
- Protozoarios 1Documento19 páginasProtozoarios 1HansPomaMolleapazaAún no hay calificaciones
- RotíferosDocumento8 páginasRotíferosFernanda Cortes50% (2)
- FILO ROTIFERA InfoDocumento5 páginasFILO ROTIFERA InfoAna MariaAún no hay calificaciones
- Biologia General - Practica N°9-F3-Diversidad de La Vida Eubacterias, Protistas y HongosDocumento14 páginasBiologia General - Practica N°9-F3-Diversidad de La Vida Eubacterias, Protistas y HongosCristian Richard Saico Ccope0% (1)
- Foraminíferos: microfósiles marinos crucialesDocumento22 páginasForaminíferos: microfósiles marinos crucialesmochavez0% (1)
- Plancton animal marinoDocumento24 páginasPlancton animal marinojustino oyono efa mangueAún no hay calificaciones
- Fico 6Documento6 páginasFico 6Jessica Yazbel Romero RicoAún no hay calificaciones
- Phylum RotíferaDocumento11 páginasPhylum RotíferaMayra BrionesAún no hay calificaciones
- Cloraracniófitos, Rhadiolarios y ForaminíferosDocumento4 páginasCloraracniófitos, Rhadiolarios y ForaminíferosAlejandraAún no hay calificaciones
- PORÍFEROSDocumento5 páginasPORÍFEROSAnonymous bt1IdYAún no hay calificaciones
- Resumen - Porifera - Las Esponjas y Su Importancia en La Biologã - A MarinaDocumento4 páginasResumen - Porifera - Las Esponjas y Su Importancia en La Biologã - A MarinaAna GarcíaAún no hay calificaciones
- ProtistaDocumento5 páginasProtistaFreymarAparicioAún no hay calificaciones
- AlgaDocumento45 páginasAlgaJESULAINAún no hay calificaciones
- Tarea Con Normas APADocumento17 páginasTarea Con Normas APAPrimeroBItscoAún no hay calificaciones
- Genaralidades PoríferosDocumento19 páginasGenaralidades PoríferosMiguel CalderonAún no hay calificaciones
- Toaz - Info Universidad Nacional Departamento Academico de San Agustin de Biologia Facultad PRDocumento336 páginasToaz - Info Universidad Nacional Departamento Academico de San Agustin de Biologia Facultad PRJOSE MACHACA CONDORIAún no hay calificaciones
- Orden Isopoda 1Documento14 páginasOrden Isopoda 1p_romero_cAún no hay calificaciones
- Protistas My BelovedDocumento3 páginasProtistas My BelovedDanny GonzalezAún no hay calificaciones
- 5 PoriferaDocumento19 páginas5 PoriferaMIGUEL ALEX PEDRAZA ARANDOAún no hay calificaciones
- 9 Filos BiologiaDocumento8 páginas9 Filos BiologiaJaime GutierrezAún no hay calificaciones
- Cetologia 1Documento4 páginasCetologia 1joelramirezlopez100% (1)
- Los 5 ReinosDocumento19 páginasLos 5 Reinosapi-260964561Aún no hay calificaciones
- Introducción a los protozoos: características, tipos y ejemplosDocumento21 páginasIntroducción a los protozoos: características, tipos y ejemplosana mariaAún no hay calificaciones
- Práctica 2. ProtozoariosDocumento9 páginasPráctica 2. ProtozoariosMayra Moreno RomeroAún no hay calificaciones
- Caceres Estrada Iacomo Fernando N°2Documento14 páginasCaceres Estrada Iacomo Fernando N°2CACERES ESTRADA IACOMO FERNANDOAún no hay calificaciones
- Importancia Ecológica y Económica de Cefalocordados y Urocordados (Esnerly Martínez y Ronaldo Reyes)Documento10 páginasImportancia Ecológica y Económica de Cefalocordados y Urocordados (Esnerly Martínez y Ronaldo Reyes)Ronaldo ReyesAún no hay calificaciones
- Hidras Hidrozoos y Anemonas 1Documento52 páginasHidras Hidrozoos y Anemonas 1DianaPoAún no hay calificaciones
- CUESTINARIO CrustaceosDocumento5 páginasCUESTINARIO CrustaceosAdriana SantosAún no hay calificaciones
- Cartilla Cnidaria PDFDocumento7 páginasCartilla Cnidaria PDFGefry Andres Castro JimenezAún no hay calificaciones
- Monografía Sobre El Orden OdonataDocumento9 páginasMonografía Sobre El Orden OdonataJean AlcantaraAún no hay calificaciones
- Práctico N°6 - Platyhelminthes y NemertiniDocumento11 páginasPráctico N°6 - Platyhelminthes y NemertiniCristian Santander RosalesAún no hay calificaciones
- La Diversidad de Los ProtistasDocumento46 páginasLa Diversidad de Los ProtistasXochitl TGAún no hay calificaciones
- PROTISTADocumento6 páginasPROTISTAazmobileguiniAún no hay calificaciones
- Informe Invertebrados Practica - 1Documento5 páginasInforme Invertebrados Practica - 1DANIEL CAMILO FLOREZ SANTANAAún no hay calificaciones
- Características Morfo-Fisiológicas en El Proceso Reproductivo de Los Seres Vivos (Animales Vertebrados)Documento16 páginasCaracterísticas Morfo-Fisiológicas en El Proceso Reproductivo de Los Seres Vivos (Animales Vertebrados)Neylic InciarteAún no hay calificaciones
- Resumen de El Proceso de Hominización: RESÚMENES UNIVERSITARIOSDe EverandResumen de El Proceso de Hominización: RESÚMENES UNIVERSITARIOSAún no hay calificaciones
- El ajolote: Biología del anfibio más sobresaliente del mundoDe EverandEl ajolote: Biología del anfibio más sobresaliente del mundoCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Catálogo de los copépodos (Crustacea: Copepoda: Calanoida y Cyclopoida) de cuerpos de agua temporales de Jalisco, México: Taxonomía y distribuciónDe EverandCatálogo de los copépodos (Crustacea: Copepoda: Calanoida y Cyclopoida) de cuerpos de agua temporales de Jalisco, México: Taxonomía y distribuciónAún no hay calificaciones
- Seminario NocheDocumento2 páginasSeminario NocheSoee CaminoAún no hay calificaciones
- CONCRETODocumento2 páginasCONCRETOSoee CaminoAún no hay calificaciones
- Constancia de PosesionDocumento1 páginaConstancia de PosesionSoee CaminoAún no hay calificaciones
- Informe de Valorizacion 001Documento1 páginaInforme de Valorizacion 001Soee CaminoAún no hay calificaciones
- Dimensión Política - EconómicaDocumento2 páginasDimensión Política - EconómicaSoee CaminoAún no hay calificaciones
- Separ AdoresDocumento29 páginasSepar AdoresSoee CaminoAún no hay calificaciones
- 4.4 Estado Situacional de La ObraDocumento2 páginas4.4 Estado Situacional de La ObraSoee CaminoAún no hay calificaciones
- Analisis Ecologico y DeficitDocumento5 páginasAnalisis Ecologico y DeficitSoee CaminoAún no hay calificaciones
- ÁncashDocumento1 páginaÁncashSoee CaminoAún no hay calificaciones
- Malecon GuayaquilDocumento16 páginasMalecon GuayaquilSoee CaminoAún no hay calificaciones
- Las Esponjas o La Porífera Ya Fueron Descritas Por Aristóteles (Recuperado Automáticamente)Documento7 páginasLas Esponjas o La Porífera Ya Fueron Descritas Por Aristóteles (Recuperado Automáticamente)Soee CaminoAún no hay calificaciones
- Mapeo y Tipología de La Expansión Urbana enDocumento10 páginasMapeo y Tipología de La Expansión Urbana enSoee CaminoAún no hay calificaciones
- Actas y Requisitos FTSDocumento26 páginasActas y Requisitos FTSSoee CaminoAún no hay calificaciones
- URBANISMODocumento4 páginasURBANISMOSoee CaminoAún no hay calificaciones
- Vision-Mision - SeguridadDocumento2 páginasVision-Mision - SeguridadSoee CaminoAún no hay calificaciones
- HABERMASDocumento4 páginasHABERMASSoee CaminoAún no hay calificaciones
- Carta 001 EspecialistaDocumento1 páginaCarta 001 EspecialistaSoee CaminoAún no hay calificaciones
- Construcción de cobertura en IE 86371 Juvenal Soto CaussoDocumento4 páginasConstrucción de cobertura en IE 86371 Juvenal Soto CaussoSoee CaminoAún no hay calificaciones
- Formato AtsDocumento1 páginaFormato AtsRicardo Poémape ChirinosAún no hay calificaciones
- PomallucayDocumento8 páginasPomallucaySoee CaminoAún no hay calificaciones
- Formato de Analisis Seguro de Trabajo ASTDocumento1 páginaFormato de Analisis Seguro de Trabajo ASTKarenAún no hay calificaciones
- Nombre de FoldersDocumento8 páginasNombre de FoldersSoee CaminoAún no hay calificaciones
- PensionDocumento1 páginaPensionSoee CaminoAún no hay calificaciones
- Sistema CostructivoDocumento20 páginasSistema CostructivoZōe CaminoAún no hay calificaciones
- Pago personal 12 julioDocumento1 páginaPago personal 12 julioSoee CaminoAún no hay calificaciones
- NORMATIVIDADDocumento14 páginasNORMATIVIDADSoee CaminoAún no hay calificaciones
- Tachos OficinaDocumento4 páginasTachos OficinaSoee CaminoAún no hay calificaciones
- UrbanismoDocumento20 páginasUrbanismoZōe CaminoAún no hay calificaciones
- UrbanismoDocumento20 páginasUrbanismoZōe CaminoAún no hay calificaciones
- Test de FrustracionDocumento22 páginasTest de FrustracionLeydi Quispe leqque100% (2)
- Bloque 2 Dosificación de Contenidos Geografia 6° GradoDocumento5 páginasBloque 2 Dosificación de Contenidos Geografia 6° Gradomasterhousez6100% (1)
- Capitulo 7 de Heterótrofos A AutótrofosDocumento3 páginasCapitulo 7 de Heterótrofos A AutótrofosSael SáezAún no hay calificaciones
- AMT 44g La Medicina Tradicional en La Sierra Centro-SurDocumento4 páginasAMT 44g La Medicina Tradicional en La Sierra Centro-SurhugodeAún no hay calificaciones
- Apuntes de Cátedra Morfología VegetalDocumento29 páginasApuntes de Cátedra Morfología VegetalEva Angulo100% (1)
- Terminos de Situaccion y DireccionDocumento5 páginasTerminos de Situaccion y DireccionManola HernandezAún no hay calificaciones
- Hormonas Adrenocorticales PDFDocumento26 páginasHormonas Adrenocorticales PDFValentina AmayaAún no hay calificaciones
- Produccion de Leche en El Trópico Con Vacas en Pastoreo RotacionalDocumento4 páginasProduccion de Leche en El Trópico Con Vacas en Pastoreo RotacionalAnita Elizabeth Layza MendiolaAún no hay calificaciones
- Guia de Aprendizaje Nro. 29 de Ciencia y Tecnologia 4to Grado. Del 29 Al 03 de DiciembreDocumento7 páginasGuia de Aprendizaje Nro. 29 de Ciencia y Tecnologia 4to Grado. Del 29 Al 03 de Diciembredasd3asAún no hay calificaciones
- Endodoncia Colaborativo IDocumento9 páginasEndodoncia Colaborativo IIvett Cristina Gonzalez FloresAún no hay calificaciones
- PeelingsDocumento28 páginasPeelingsIrina KucevichuteAún no hay calificaciones
- La expiación: celos y secretos entre amigosDocumento22 páginasLa expiación: celos y secretos entre amigosterefAún no hay calificaciones
- fruticultura e interceptación de radiación solar en árboles frutalesDocumento5 páginasfruticultura e interceptación de radiación solar en árboles frutalesLaura MoraAún no hay calificaciones
- Tesis Suplementacion Con Harina de Moringa y Su Efecto en Conejos DestetadosDocumento68 páginasTesis Suplementacion Con Harina de Moringa y Su Efecto en Conejos DestetadosJoseRamonTraspuestoAún no hay calificaciones
- Planeación de Clase Grado SegundoDocumento6 páginasPlaneación de Clase Grado SegundoGLORIA CECILIA GARCIA GARCIAAún no hay calificaciones
- Capítulo 12 - Extremidades InformeeDocumento7 páginasCapítulo 12 - Extremidades InformeeAnonymous qA5NIMQBaAún no hay calificaciones
- Cerdo WikipediaDocumento10 páginasCerdo WikipediaTeffa Lozano SAún no hay calificaciones
- Motricidad Fina Hojas de Trabajo PreescolarDocumento23 páginasMotricidad Fina Hojas de Trabajo PreescolarLep Julian Gonzalez BarbosaAún no hay calificaciones
- Anatomia Prostatica Via AnteriorDocumento48 páginasAnatomia Prostatica Via AnteriorFelipe Rodríguez MorillaAún no hay calificaciones
- Ejercicios Prácticos para Fijar La AtenciónDocumento2 páginasEjercicios Prácticos para Fijar La AtenciónDenisse Reyes GAún no hay calificaciones
- Umbrales SensorialesDocumento9 páginasUmbrales SensorialesDelia AlcántaraAún no hay calificaciones
- Apuntes Estratigrafia Sedimentacion Emnc 1 2017Documento232 páginasApuntes Estratigrafia Sedimentacion Emnc 1 2017Sergio Santiago Rafailo100% (1)
- Trabajo Nro 3Documento29 páginasTrabajo Nro 3Arven CastilloAún no hay calificaciones
- Control de Damping off en arveja con Trichoderma harzianumDocumento29 páginasControl de Damping off en arveja con Trichoderma harzianumandrewn21Aún no hay calificaciones
- Sistema EsqueléticoDocumento3 páginasSistema EsqueléticoSandra Jacqueline Peña ValdésAún no hay calificaciones
- Levantado de TextoDocumento5 páginasLevantado de TextoLeslin Reyes MayénAún no hay calificaciones
- Laboratorio de Bioquimica LecheDocumento5 páginasLaboratorio de Bioquimica LechevictorAún no hay calificaciones
- MM Zany Octubre 4° PDFDocumento69 páginasMM Zany Octubre 4° PDFCarmen PerezAún no hay calificaciones
- Trabajo Final de Condicionamiento ClasicoDocumento10 páginasTrabajo Final de Condicionamiento ClasicoElida Espinal Martinez100% (3)
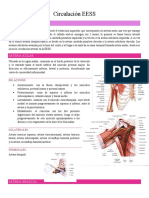
- Circulacion EESSDocumento8 páginasCirculacion EESSmelisaAún no hay calificaciones