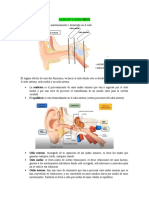
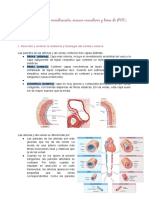
También podría gustarte
- 26 Posturas Del Bikram YogaDocumento5 páginas26 Posturas Del Bikram Yogabloodyblack100% (3)
- Trabajo Práctico 1 F de La ElectrónicaDocumento23 páginasTrabajo Práctico 1 F de La ElectrónicaMirian Yolanda GonzalezAún no hay calificaciones
- Tercer Mes Al Nacimiento - PlacentaDocumento66 páginasTercer Mes Al Nacimiento - PlacentaJessica Daniela Salazar Castaneda100% (2)
- Resumen GastrulacionDocumento2 páginasResumen Gastrulacionfireball764100% (1)
- Sanidad y Nutrición Crecimiento y Engorde Del CerdoDocumento58 páginasSanidad y Nutrición Crecimiento y Engorde Del CerdoKeyla CabreraAún no hay calificaciones
- Primera Semana Del Desarrollo HumanoDocumento3 páginasPrimera Semana Del Desarrollo HumanoMedicina UiC100% (2)
- 11° Semana MORFO II GUIA PRACT LAB FISIOL-2019Documento20 páginas11° Semana MORFO II GUIA PRACT LAB FISIOL-2019Angie VallejosAún no hay calificaciones
- División CelularDocumento10 páginasDivisión CelularAndrea Fernanda Calderon CuadrosAún no hay calificaciones
- Embriologia Abril 2015Documento109 páginasEmbriologia Abril 2015Laura Tatiana Padilla PinzonAún no hay calificaciones
- TERMORREGULACIÓNDocumento27 páginasTERMORREGULACIÓNPedro Estrella ParianAún no hay calificaciones
- Crecimiento y Desarrollo Embrionario y FetalDocumento43 páginasCrecimiento y Desarrollo Embrionario y FetalJimmy Hoffman MarleyAún no hay calificaciones
- Embriologia de LagmanDocumento5 páginasEmbriologia de LagmanMaria MoraAún no hay calificaciones
- Embriología del aparato respiratorioDocumento3 páginasEmbriología del aparato respiratorioDracula Vlad100% (1)
- Esqueleto AxialDocumento10 páginasEsqueleto AxialJcamino100% (1)
- Casoclinico AnencefaliaDocumento9 páginasCasoclinico AnencefaliaJesus Smaikel Teran MuenalaAún no hay calificaciones
- EmbriologiaDocumento5 páginasEmbriologiaJhey Kenider Benites IzquierdoAún no hay calificaciones
- Mesodermo y MesenquimaDocumento3 páginasMesodermo y MesenquimaAntoAlvaradoAún no hay calificaciones
- Capitulo 1 - Organización Funcional Del Cuerpo Humano y Control Del Medio InternoDocumento4 páginasCapitulo 1 - Organización Funcional Del Cuerpo Humano y Control Del Medio InternoMirianCamposAún no hay calificaciones
- Formación capas embrionariasDocumento2 páginasFormación capas embrionariasomarAún no hay calificaciones
- AdipocitosDocumento19 páginasAdipocitosAlejandro CampusintrifosfatoAún no hay calificaciones
- Anatomía de La YeguaDocumento15 páginasAnatomía de La YeguaPaula LealAún no hay calificaciones
- Anatomía y Embriología AnimalDocumento9 páginasAnatomía y Embriología Animalclarimar vasquez cese100% (1)
- Embriología cavidadesDocumento19 páginasEmbriología cavidadesDavenia SaldañaAún no hay calificaciones
- Glicoproteinas 2006Documento33 páginasGlicoproteinas 2006api-3708303100% (1)
- EmbriologíaDocumento17 páginasEmbriologíaJames Russo VelásquezAún no hay calificaciones
- Función y Anatomia Del OvarioDocumento4 páginasFunción y Anatomia Del OvarioMarlina TorretaAún no hay calificaciones
- Formación del disco embrionario bilaminar en la segunda semana de gestaciónDocumento34 páginasFormación del disco embrionario bilaminar en la segunda semana de gestaciónJair ReyesAún no hay calificaciones
- Resumen Anatomía Aplicada Cráneo y ExtremidadesDocumento6 páginasResumen Anatomía Aplicada Cráneo y Extremidadesanatomia_aplicada4100% (2)
- Adaptaciones Al Frío de Endotermos y EctotermosDocumento3 páginasAdaptaciones Al Frío de Endotermos y EctotermosStefanía Sibille0% (1)
- Sistema Cardiovascular. EMBRIOLOGIA PDFDocumento10 páginasSistema Cardiovascular. EMBRIOLOGIA PDFDavid De La RosaAún no hay calificaciones
- Tercera-Octava Semana de Desarrollo EmbrionarioDocumento5 páginasTercera-Octava Semana de Desarrollo Embrionarioveronica100% (1)
- Mola Hidatiforme. Seminario 6 FisiologíaDocumento42 páginasMola Hidatiforme. Seminario 6 FisiologíaJosé Roberto Sánchez Méndez100% (1)
- Embriologia PulmonarDocumento8 páginasEmbriologia PulmonarLina María CañónAún no hay calificaciones
- Cuarto ventrículo y sus estructurasDocumento9 páginasCuarto ventrículo y sus estructuraspaolo mendezAún no hay calificaciones
- DIFUSIÓNDocumento1 páginaDIFUSIÓNAdriana Olivares100% (1)
- EspermatogenesisDocumento3 páginasEspermatogenesisnumelys mendoza100% (1)
- Periodo FetalDocumento76 páginasPeriodo FetalSusan HarlemAún no hay calificaciones
- Trabajo de Investigacion HistologiaDocumento4 páginasTrabajo de Investigacion HistologiaKerluis Solorzano0% (1)
- Maduración, Plegamiento y Degradacion de Proteinas.Documento3 páginasMaduración, Plegamiento y Degradacion de Proteinas.Natalia EstradaAún no hay calificaciones
- Histología Del OídoDocumento11 páginasHistología Del OídoluisAún no hay calificaciones
- Resumen Fisiología Aparato RespiratorioDocumento20 páginasResumen Fisiología Aparato RespiratorioConni CastroAún no hay calificaciones
- Microscopia ConfocalDocumento9 páginasMicroscopia ConfocalJhordan Ojeda GonzalezAún no hay calificaciones
- Desarrollo Del Tubo Intestinal, Páncreas y DiafragmaDocumento6 páginasDesarrollo Del Tubo Intestinal, Páncreas y Diafragmaeliana bustamante0% (1)
- Bioenergética PDFDocumento6 páginasBioenergética PDFalexmaras95100% (1)
- Fisiologia Respiratoria OriginalDocumento48 páginasFisiologia Respiratoria OriginalWilliam DazaAún no hay calificaciones
- Sensibilidades Somaticas-Dolor-Cefalea y Sensibilidad TermicaDocumento3 páginasSensibilidades Somaticas-Dolor-Cefalea y Sensibilidad TermicaGmer19100% (1)
- 1 Neurulación y Creta NeuralDocumento3 páginas1 Neurulación y Creta NeuralDawdunoAún no hay calificaciones
- Tejido Cartilaginoso HistologiaDocumento18 páginasTejido Cartilaginoso HistologiaMilkaLopintaAún no hay calificaciones
- Desarrollo Embrionario Temprano 2009Documento16 páginasDesarrollo Embrionario Temprano 2009Estudiantes de Veterinaria Primer año 2009Aún no hay calificaciones
- 1 Cavidades Corporales y OsteomuscularDocumento48 páginas1 Cavidades Corporales y OsteomuscularShalom ApérrigueAún no hay calificaciones
- Anatomia Del BazoDocumento3 páginasAnatomia Del BazoClaudia L. PerezAún no hay calificaciones
- Presentación El Sistema Reproductor Masculino y FemeninoDocumento15 páginasPresentación El Sistema Reproductor Masculino y FemeninoeliasAún no hay calificaciones
- Transporte de GametosDocumento12 páginasTransporte de GametosPerla MorenoAún no hay calificaciones
- E1 - Ciclo Ovarico y Endometrial 2017Documento10 páginasE1 - Ciclo Ovarico y Endometrial 2017Marcelino GaravitoAún no hay calificaciones
- ProtuberanciaDocumento7 páginasProtuberanciaSteven JiménezAún no hay calificaciones
- Vascularizacion Del CuelloDocumento21 páginasVascularizacion Del CuelloJJ AnchundiaAún no hay calificaciones
- 7.2) El Cromosoma - Organizacion Del ADNDocumento45 páginas7.2) El Cromosoma - Organizacion Del ADNSzzczzAún no hay calificaciones
- Epitelios y Clasificación de GlándulasDocumento44 páginasEpitelios y Clasificación de GlándulasHenry Petit RodriguezAún no hay calificaciones
- Desarrollo Del Aparato RespiratorioDocumento2 páginasDesarrollo Del Aparato RespiratorioAngel Stevens Garcia CallaoAún no hay calificaciones
- AsquelmintosDocumento4 páginasAsquelmintosDavidan1022Aún no hay calificaciones
- Abdomen de Los InsectosDocumento13 páginasAbdomen de Los InsectosAnthony Roberto Cerdán Jimenez100% (2)
- Etapas Del Desarrollo Embrionario en AnfibiosDocumento4 páginasEtapas Del Desarrollo Embrionario en AnfibiosOlinto Vicente Benedetti CarpioAún no hay calificaciones
- 01 CruzJos-TerapiaCCenelmanejodelduelocomplicado PDFDocumento20 páginas01 CruzJos-TerapiaCCenelmanejodelduelocomplicado PDFLuz duranAún no hay calificaciones
- MOLUSCOSDocumento1 páginaMOLUSCOSabigail juarezAún no hay calificaciones
- La Biologia en La IudadDocumento1 páginaLa Biologia en La Iudadabigail juarezAún no hay calificaciones
- MOLUSCOSDocumento1 páginaMOLUSCOSabigail juarezAún no hay calificaciones
- EVOLUCIONDocumento3 páginasEVOLUCIONabigail juarezAún no hay calificaciones
- ANATOMIADocumento3 páginasANATOMIAabigail juarezAún no hay calificaciones
- ORGANISMOSDocumento2 páginasORGANISMOSabigail juarezAún no hay calificaciones
- BIOLEMENTOSDocumento4 páginasBIOLEMENTOSabigail juarezAún no hay calificaciones
- REFERENCIASDocumento1 páginaREFERENCIASabigail juarezAún no hay calificaciones
- Farmacologia GeneralDocumento214 páginasFarmacologia GeneralCarolinalcmAún no hay calificaciones
- INTRODUCCIONDocumento3 páginasINTRODUCCIONabigail juarezAún no hay calificaciones
- REFERENCIASDocumento1 páginaREFERENCIASabigail juarezAún no hay calificaciones
- HISTORIADocumento4 páginasHISTORIAabigail juarezAún no hay calificaciones
- COMPONENTESDocumento8 páginasCOMPONENTESabigail juarezAún no hay calificaciones
- Medidas de Prevención de Contagio en La EmpresaDocumento3 páginasMedidas de Prevención de Contagio en La EmpresaIUSMOAún no hay calificaciones
- Promoción de La Salud PDFDocumento1 páginaPromoción de La Salud PDFlucho rojasAún no hay calificaciones
- Armacología en EnfermeríaDocumento1 páginaArmacología en Enfermeríaabigail juarezAún no hay calificaciones
- Cartel Nom 030 Stps 2008Documento1 páginaCartel Nom 030 Stps 2008abigail juarezAún no hay calificaciones
- Guia de Medicamentos Esenciales ComprimidoDocumento153 páginasGuia de Medicamentos Esenciales ComprimidoGüids GalliganiAún no hay calificaciones
- COMPETENCIASDocumento1 páginaCOMPETENCIASabigail juarezAún no hay calificaciones
- Farmacologia GeneralDocumento214 páginasFarmacologia GeneralCarolinalcmAún no hay calificaciones
- Manual de Practicas de Farmacologia Clinica - Enfermeria ORNELAS-UNAM 2016Documento133 páginasManual de Practicas de Farmacologia Clinica - Enfermeria ORNELAS-UNAM 2016javier chavez100% (3)
- El Conocimiento de La Farmacología en El Profesional de EnfermeríaDocumento10 páginasEl Conocimiento de La Farmacología en El Profesional de EnfermeríaJj CpAún no hay calificaciones
- AGRUPACIONDocumento2 páginasAGRUPACIONabigail juarezAún no hay calificaciones
- La ProduccionDocumento5 páginasLa Produccionabigail juarezAún no hay calificaciones
- BandaDocumento2 páginasBandaabigail juarezAún no hay calificaciones
- Manual de Practicas de Farmacologia Clinica - Enfermeria ORNELAS-UNAM 2016Documento133 páginasManual de Practicas de Farmacologia Clinica - Enfermeria ORNELAS-UNAM 2016javier chavez100% (3)
- Segunda OlaDocumento3 páginasSegunda Olaabigail juarezAún no hay calificaciones
- Mijito RioDocumento1 páginaMijito Rioabigail juarezAún no hay calificaciones
- Ego Caso 1Documento2 páginasEgo Caso 1Cruz AlonsoAún no hay calificaciones
- Anatomía y FisiologíaDocumento64 páginasAnatomía y FisiologíaantoniobitchAún no hay calificaciones
- Tríptico. Mesón 2.Documento2 páginasTríptico. Mesón 2.Diana LugoAún no hay calificaciones
- Estudio de La Actividad Aminopeptidasica en Espermatozoides Astenozoospermicos. Comparacion Clinica PDFDocumento249 páginasEstudio de La Actividad Aminopeptidasica en Espermatozoides Astenozoospermicos. Comparacion Clinica PDFAco ChapaAún no hay calificaciones
- Generalidades Del CorazónDocumento11 páginasGeneralidades Del CorazónAntu BelmaresAún no hay calificaciones
- Motilidad Celular - LisDocumento3 páginasMotilidad Celular - LisDEBORA LISBET ROBLES TUESTAAún no hay calificaciones
- Células Del Sistema Inmunitario y Fisiología Del Sistema InmuneDocumento8 páginasCélulas Del Sistema Inmunitario y Fisiología Del Sistema InmuneJulio RodríguezAún no hay calificaciones
- Taller de Sistema Linfatico # 1Documento6 páginasTaller de Sistema Linfatico # 1Thiago GarciaAún no hay calificaciones
- Fisio NeurologíaDocumento7 páginasFisio NeurologíaJUAN DIEGO CUSIPAUCAR CARDENASAún no hay calificaciones
- Resumen de Glandula SuprarrenalDocumento5 páginasResumen de Glandula SuprarrenalFuentes Chevez RubenAún no hay calificaciones
- Admin,+art +6 1Documento33 páginasAdmin,+art +6 1IZAMAR NADDINE LLAURE FLORESAún no hay calificaciones
- Guía N°2 - Ciclo Celular - MITOSIS - II-medioDocumento9 páginasGuía N°2 - Ciclo Celular - MITOSIS - II-medioregulus blackAún no hay calificaciones
- PatoDocumento8 páginasPatoDanixaKristelBarriosUcañayAún no hay calificaciones
- Anatomia Del OidoDocumento47 páginasAnatomia Del OidoAndrea NarvaezAún no hay calificaciones
- Curso Terapia Puntos MarmaDocumento46 páginasCurso Terapia Puntos MarmaCatherine MaechtigAún no hay calificaciones
- Microscopia Y Técnica HistologicaDocumento27 páginasMicroscopia Y Técnica HistologicaJeffer RomeroAún no hay calificaciones
- Programa Del Curso de Entrenador Territorial de BalonmanoDocumento20 páginasPrograma Del Curso de Entrenador Territorial de BalonmanoOtsenreAún no hay calificaciones
- Taller 7 Los TejidosDocumento4 páginasTaller 7 Los TejidosOmaira TorresAún no hay calificaciones
- Tema 3 Artrolog 1 ParteDocumento56 páginasTema 3 Artrolog 1 ParteRobeccaAún no hay calificaciones
- Punción Subclavia, Venodisección, Accesos Vasculares y Toma de PVC.Documento33 páginasPunción Subclavia, Venodisección, Accesos Vasculares y Toma de PVC.MANCILLA RODRIGUEZ GRECIA NAZARETAún no hay calificaciones
- Preguntas Sistema Reproductor MasculinoDocumento4 páginasPreguntas Sistema Reproductor MasculinoCatalina Andrea Soto100% (5)
- Anatomia 2doDocumento4 páginasAnatomia 2doCielo Martinez LopezAún no hay calificaciones
- Anastomosis Porto-CavaDocumento2 páginasAnastomosis Porto-CavaPau Rodriguez100% (1)
- Biologia y Anatomia - Volumen IiDocumento14 páginasBiologia y Anatomia - Volumen IiBrazhia DueñasAún no hay calificaciones
- Tema 3 Junto Wuolah VozDocumento9 páginasTema 3 Junto Wuolah VozMaría García-chicoAún no hay calificaciones
- Sistema ExcretorDocumento3 páginasSistema ExcretorDiana Isabel Ochoa YanezAún no hay calificaciones
- Desarrollo de La Columna VertebralDocumento4 páginasDesarrollo de La Columna VertebralRosmary HornaAún no hay calificaciones
- Sistema CirculatorioDocumento27 páginasSistema CirculatorioNeymar MamaniAún no hay calificaciones