También podría gustarte
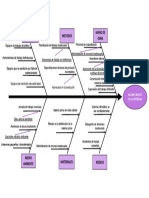
- Diagrama Ishikawa 4Documento1 páginaDiagrama Ishikawa 4Mabel AllsAún no hay calificaciones
- La energía y la revolución industrialDocumento5 páginasLa energía y la revolución industrialEder Aranda CarbonellAún no hay calificaciones
- Value Proposition CanvasDocumento1 páginaValue Proposition CanvasJuan Pablo LabraAún no hay calificaciones
- Evaluación Continua 2Documento3 páginasEvaluación Continua 2Giselle Torres LopezAún no hay calificaciones
- Apuntes de Antenas PDFDocumento84 páginasApuntes de Antenas PDFSaul De Leon EstradaAún no hay calificaciones
- Estabilización Con GeosinteticosDocumento106 páginasEstabilización Con GeosinteticosEdward Alayo100% (1)
- Gestion de Residuos SólidosDocumento19 páginasGestion de Residuos SólidosGiselle Torres LopezAún no hay calificaciones
- Evidencia AA2 - Ev2 - Informe Planeación Del Soporte TécnicoDocumento3 páginasEvidencia AA2 - Ev2 - Informe Planeación Del Soporte TécnicoAlex Loboa Cortes50% (8)
- ENRNC Biomasa en ChileDocumento22 páginasENRNC Biomasa en ChiledanielAún no hay calificaciones
- Documento de Vibraciones PDFDocumento114 páginasDocumento de Vibraciones PDFGeorAún no hay calificaciones
- Apuntes Basicos de Automatizacion Ing. Eddy PerezDocumento53 páginasApuntes Basicos de Automatizacion Ing. Eddy PerezeddyAún no hay calificaciones
- Absorcion y Transporte Del Cadmio Por Las Plantas Grupo HuancaDocumento7 páginasAbsorcion y Transporte Del Cadmio Por Las Plantas Grupo HuancareymundohuancaAún no hay calificaciones
- Plan de Mercadeo para artesanos de Baños del IncaDocumento7 páginasPlan de Mercadeo para artesanos de Baños del Incamasterbaldomero2012Aún no hay calificaciones
- Biotecnologia MicrobianaDocumento14 páginasBiotecnologia MicrobianaJose david Orozco avilaAún no hay calificaciones
- Monografía Comercio ExteriorDocumento10 páginasMonografía Comercio Exteriormaria olivera alzamoraAún no hay calificaciones
- Programas de Protección Regional de Áreas Prioritarias de grandes ballenas en el golfo de California y costa occidental de Baja California Sur: Propuestas de conservaciónDe EverandProgramas de Protección Regional de Áreas Prioritarias de grandes ballenas en el golfo de California y costa occidental de Baja California Sur: Propuestas de conservaciónAún no hay calificaciones
- Contaminación Por FertilizantesDocumento14 páginasContaminación Por FertilizanteskakarottoflyAún no hay calificaciones
- Cinética de Crecimiento MicrobianoDocumento6 páginasCinética de Crecimiento MicrobianoJuan Montalvo CerronAún no hay calificaciones
- Hidrógeno Como Combustible AlternativoDocumento65 páginasHidrógeno Como Combustible AlternativopoliticeanAún no hay calificaciones
- Marco TeóricoDocumento3 páginasMarco TeóricoDominikq123Aún no hay calificaciones
- Producción de Biodiesel A Partir de Aceite de PalmaDocumento4 páginasProducción de Biodiesel A Partir de Aceite de PalmaDario Fernando Yepez Vela100% (1)
- Deinococcus RadioduransDocumento10 páginasDeinococcus RadioduransPaula AngaritaAún no hay calificaciones
- E.hernandez Reflexion - FotosintesisDocumento5 páginasE.hernandez Reflexion - FotosintesisEstefania HernandezAún no hay calificaciones
- Reformado Con VaporDocumento10 páginasReformado Con VaporDiego FernandoAún no hay calificaciones
- El Agua PotableDocumento154 páginasEl Agua PotableArturo Othoniel Mendoza Marín100% (1)
- Identificación y reacciones de alcoholes orgánicosDocumento7 páginasIdentificación y reacciones de alcoholes orgánicosRuth Ojeda DominguezAún no hay calificaciones
- El HidrogenoDocumento20 páginasEl HidrogenoLaura Salcedo FladineAún no hay calificaciones
- Proyecto Final Jorge GarciaDocumento24 páginasProyecto Final Jorge GarciayogeunysAún no hay calificaciones
- Jefferson Fernandez - QuimicaDocumento7 páginasJefferson Fernandez - QuimicaJefry FernandezAún no hay calificaciones
- Monografico de Socio LA NIEBLA de LONDRESDocumento16 páginasMonografico de Socio LA NIEBLA de LONDRESLizeth SukAún no hay calificaciones
- Guai No 4 Determinación Dureza en Agua, Huemdad y CenizasDocumento9 páginasGuai No 4 Determinación Dureza en Agua, Huemdad y CenizasCamilo MartínezAún no hay calificaciones
- Tecnologia EnzimaticaDocumento7 páginasTecnologia EnzimaticaErandeny Medina Morales-Simmons100% (6)
- Deber de Biología RESUELTO - Bachillerato - Proyecto3 - Semana 1Documento4 páginasDeber de Biología RESUELTO - Bachillerato - Proyecto3 - Semana 1Otto Franco SotomayorAún no hay calificaciones
- Coloides Del SueloDocumento12 páginasColoides Del SueloCielo Moreno100% (2)
- Transferencia de momento lineal en sistemas biológicosDocumento30 páginasTransferencia de momento lineal en sistemas biológicosAguilar Alvarado Josué AarónAún no hay calificaciones
- Evolución de La BiotecnologíaDocumento9 páginasEvolución de La BiotecnologíaJoeKilverCornejoSandovalAún no hay calificaciones
- El ColagenoDocumento4 páginasEl ColagenoYohanny CastroAún no hay calificaciones
- Sustentabilidad EcologicaDocumento5 páginasSustentabilidad EcologicaFrancisco Altamirano100% (1)
- FotofosforilacionDocumento26 páginasFotofosforilacionChristian Leo RuizAún no hay calificaciones
- Smog FotoquimicoDocumento4 páginasSmog FotoquimicoBranco Figueroa AyalaAún no hay calificaciones
- LOMBRIHUMUSDocumento58 páginasLOMBRIHUMUSELIZABETHAún no hay calificaciones
- Sem1.conceptos Básicos de Ingeniería AmbientalDocumento43 páginasSem1.conceptos Básicos de Ingeniería AmbientalAndreita OrTssAún no hay calificaciones
- INTRODUCCIÓN de Biogas ProyectoDocumento83 páginasINTRODUCCIÓN de Biogas ProyectoDiony Calloapaza100% (1)
- Ciclo contaminaciónDocumento6 páginasCiclo contaminaciónElizabeth Torres Miranda50% (2)
- Plaguicidas PanamáDocumento6 páginasPlaguicidas PanamáDidimo100% (1)
- Analisis Ciclo de Vida PDFDocumento32 páginasAnalisis Ciclo de Vida PDFLuciana Pachon0% (1)
- Fermentacion AlcoholicaDocumento3 páginasFermentacion Alcoholicapako7_15Aún no hay calificaciones
- Poliester Exposicion ResumenDocumento4 páginasPoliester Exposicion ResumenAnonymous kBHntTVYAún no hay calificaciones
- Catálisis BifásicaDocumento8 páginasCatálisis BifásicaGénesis FalcónAún no hay calificaciones
- Convenio de Estocolmo Sobre Contaminantes Orgánicos Persistentes. Estocolmo, 22 Mayo 2001Documento41 páginasConvenio de Estocolmo Sobre Contaminantes Orgánicos Persistentes. Estocolmo, 22 Mayo 2001dipublico100% (1)
- Qué Es Ciclo Del NitrógenoDocumento3 páginasQué Es Ciclo Del NitrógenoMari ChrAún no hay calificaciones
- Práctica #9 Estereoquímica Epiam 2021 ADocumento13 páginasPráctica #9 Estereoquímica Epiam 2021 AZENAIDA MADELEY PRUDENCIO QUISPEAún no hay calificaciones
- Bioetanol de Caña de AzucarDocumento28 páginasBioetanol de Caña de AzucarLAURA LUC�A ATENCIA CASTILLOAún no hay calificaciones
- Determinación de La Tasa Fotosintética de Una PlantaDocumento3 páginasDeterminación de La Tasa Fotosintética de Una PlantaSantiago Quiroga TAún no hay calificaciones
- BiogasDocumento50 páginasBiogasMari LizAún no hay calificaciones
- Huella de CarbonoDocumento24 páginasHuella de CarbonoSócrates Fernández KucharczykAún no hay calificaciones
- 2.5. Transformaciones BiológicasDocumento12 páginas2.5. Transformaciones BiológicasEmmaColladoArredondoAún no hay calificaciones
- Toxicidad del aluminio en suelos ácidosDocumento10 páginasToxicidad del aluminio en suelos ácidosPedro Benavente DonayreAún no hay calificaciones
- Biodegradabilidad de DetergentesDocumento35 páginasBiodegradabilidad de DetergentesEdson Yuri CavidesAún no hay calificaciones
- Clasificacion de Los Cuerpos Hidricos Segun La Ana Contaminacion de Agu CorregidoasDocumento54 páginasClasificacion de Los Cuerpos Hidricos Segun La Ana Contaminacion de Agu CorregidoasKarina Aycache100% (1)
- Reacciones AnódicasDocumento4 páginasReacciones AnódicasYanina DíazAún no hay calificaciones
- Biofísica lípidosDocumento4 páginasBiofísica lípidosSamuel Aldair50% (2)
- Reacciones de La Fase Luminosa de La FotosíntesisDocumento3 páginasReacciones de La Fase Luminosa de La FotosíntesisCarmen AvilaAún no hay calificaciones
- Informe de Laboratorio BioquimicaDocumento11 páginasInforme de Laboratorio BioquimicaChuCho GonzalezAún no hay calificaciones
- Biocombustibles EnsayoDocumento11 páginasBiocombustibles Ensayoanon_501722255Aún no hay calificaciones
- UVMProcBiol3erExPar2019 02Documento2 páginasUVMProcBiol3erExPar2019 02José Francisco Macias OrtegaAún no hay calificaciones
- Competencia Microbiana y CooperaciónDocumento4 páginasCompetencia Microbiana y CooperaciónFernando GarciaAún no hay calificaciones
- Dqo STM 5220 DDocumento38 páginasDqo STM 5220 Dmak gonzalesAún no hay calificaciones
- Experimento papel, agua y fuegoDocumento3 páginasExperimento papel, agua y fuegoDianaFonsecaAún no hay calificaciones
- 2° Practico InformaticaDocumento10 páginas2° Practico InformaticaWilson Lobo Davila ZAún no hay calificaciones
- Ciclos BiogeoquimicosDocumento10 páginasCiclos BiogeoquimicosedgarAún no hay calificaciones
- Formula RioDocumento2 páginasFormula RioGiselle Torres LopezAún no hay calificaciones
- Balance Hidrico Grupo 3Documento15 páginasBalance Hidrico Grupo 3Giselle Torres LopezAún no hay calificaciones
- Reforma vestuarios personal IFEMADocumento153 páginasReforma vestuarios personal IFEMAGiselle Torres LopezAún no hay calificaciones
- Reading ComprenhensionDocumento5 páginasReading ComprenhensionGiselle Torres LopezAún no hay calificaciones
- Triptico Cuencas Grupo 3Documento2 páginasTriptico Cuencas Grupo 3Giselle Torres LopezAún no hay calificaciones
- Anexo 2.4 Formato RecursoDocumento1 páginaAnexo 2.4 Formato RecursoGiselle Torres LopezAún no hay calificaciones
- Balance Hidrico-Est CanchacallaDocumento142 páginasBalance Hidrico-Est CanchacallaGiselle Torres LopezAún no hay calificaciones
- Módulo 7Documento12 páginasMódulo 7Giselle Torres LopezAún no hay calificaciones
- PROBLEMA 2: OD vs distancia con y sin vertidoDocumento3 páginasPROBLEMA 2: OD vs distancia con y sin vertidoGiselle Torres LopezAún no hay calificaciones
- Estación de Transferencia (GRUPO 3) Administrativo TallerDocumento1 páginaEstación de Transferencia (GRUPO 3) Administrativo TallerGiselle Torres LopezAún no hay calificaciones
- Record AcademicoDocumento2 páginasRecord AcademicoGiselle Torres LopezAún no hay calificaciones
- Practica Calificada 1Documento2 páginasPractica Calificada 1Giselle Torres LopezAún no hay calificaciones
- Ingenieria EconomicaDocumento10 páginasIngenieria EconomicaGiselle Torres LopezAún no hay calificaciones
- Problema 1Documento2 páginasProblema 1Giselle Torres LopezAún no hay calificaciones
- Taller 5Documento10 páginasTaller 5Giselle Torres LopezAún no hay calificaciones
- Problema 3 (Torres Lopez Giselle)Documento2 páginasProblema 3 (Torres Lopez Giselle)Giselle Torres LopezAún no hay calificaciones
- ImportanciaDocumento5 páginasImportanciaGiselle Torres LopezAún no hay calificaciones
- RESIDUOSDocumento4 páginasRESIDUOSGiselle Torres LopezAún no hay calificaciones
- Semana NDocumento4 páginasSemana NGiselle Torres LopezAún no hay calificaciones
- Objetiv OsDocumento1 páginaObjetiv OsGiselle Torres LopezAún no hay calificaciones
- So 2Documento2 páginasSo 2Giselle Torres LopezAún no hay calificaciones
- SeguridadDocumento3 páginasSeguridadGiselle Torres LopezAún no hay calificaciones
- SeguridadDocumento3 páginasSeguridadGiselle Torres LopezAún no hay calificaciones
- Semana NDocumento2 páginasSemana NGiselle Torres LopezAún no hay calificaciones
- Correción de Examen ParcialDocumento5 páginasCorreción de Examen ParcialGiselle Torres LopezAún no hay calificaciones
- Vacíos LegalesDocumento3 páginasVacíos LegalesAntonio RamirezAún no hay calificaciones
- Formato SolicitudDocumento3 páginasFormato SolicitudricardoAún no hay calificaciones
- UnacDocumento5 páginasUnacGiselle Torres LopezAún no hay calificaciones
- AEREOTRIANGULACIONDocumento4 páginasAEREOTRIANGULACIONDaniel Arando ChoqueAún no hay calificaciones
- Expo Rio PanceDocumento6 páginasExpo Rio PanceLaura ValenciaAún no hay calificaciones
- Modelo ER universidadDocumento15 páginasModelo ER universidadangelica rodriguezAún no hay calificaciones
- Practica13 Gauss-JordanDocumento4 páginasPractica13 Gauss-JordanSalvador DuranAún no hay calificaciones
- 2020 II Tema 3 Ec 511 HDocumento48 páginas2020 II Tema 3 Ec 511 HMANUEL ANTONIO BENITES RAMIREZAún no hay calificaciones
- Fichas bibliográficas sobre el uso de recursos audiovisuales en la comprensión lectoraDocumento4 páginasFichas bibliográficas sobre el uso de recursos audiovisuales en la comprensión lectoraUlices SalasAún no hay calificaciones
- Caso de Estudio Fruta FresaDocumento7 páginasCaso de Estudio Fruta FresaLizeth DiazAún no hay calificaciones
- TripodeDocumento7 páginasTripodenancitaAún no hay calificaciones
- Formación Dual para Técnico en Desabolladura de VehículosDocumento5 páginasFormación Dual para Técnico en Desabolladura de VehículosNaruto UzumakiAún no hay calificaciones
- Problema - Regla de ChvorinovDocumento3 páginasProblema - Regla de ChvorinovRaul AlcántaraAún no hay calificaciones
- Ecosistemas Acuáticos de ColombiaDocumento9 páginasEcosistemas Acuáticos de ColombiaangeloAún no hay calificaciones
- Manten Tu Colegio Limpio TallerDocumento4 páginasManten Tu Colegio Limpio TallerCarlosAún no hay calificaciones
- Historia ASCIIDocumento5 páginasHistoria ASCIICesar PanjojAún no hay calificaciones
- Conversor de Lumen A LuxDocumento1 páginaConversor de Lumen A LuxCarlos QuirozAún no hay calificaciones
- ReciboEntelMarzoDocumento2 páginasReciboEntelMarzoGeraldine GeraldineAún no hay calificaciones
- El Tiempo y El EspacioDocumento4 páginasEl Tiempo y El Espaciorochu tempriAún no hay calificaciones
- 8 Tesis - Impacto Psicosocial y Calidad de Vida en Mujeres Con Endometriosis - Situación Actual en España PDFDocumento110 páginas8 Tesis - Impacto Psicosocial y Calidad de Vida en Mujeres Con Endometriosis - Situación Actual en España PDFAzucenaAún no hay calificaciones
- Moncada 2018. Agrosofía Saberes CampesinosDocumento104 páginasMoncada 2018. Agrosofía Saberes CampesinosCorrecciones DianaAún no hay calificaciones
- INTENTO 2 Parcial - Escenario 4 - SEGUNDO BLOQUE-TEORICO - PRACTICO - ERGONOMIA - (GRUPO B02)Documento9 páginasINTENTO 2 Parcial - Escenario 4 - SEGUNDO BLOQUE-TEORICO - PRACTICO - ERGONOMIA - (GRUPO B02)verónica DíazAún no hay calificaciones
- Manual Consumos Producciones PDFDocumento7 páginasManual Consumos Producciones PDFMaria Eugenia ValeraAún no hay calificaciones
- Modelo NPZD FitoPlanctonDocumento49 páginasModelo NPZD FitoPlanctonFany Monterrubio LozanoAún no hay calificaciones