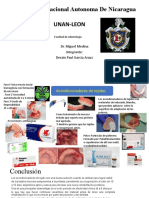
También podría gustarte
- Atencion A La NiñezDocumento11 páginasAtencion A La NiñezPaul GarciaAún no hay calificaciones
- Cronograma de TrabajoDocumento1 páginaCronograma de TrabajoPaul GarciaAún no hay calificaciones
- GingivectomíaDocumento9 páginasGingivectomíaPaul GarciaAún no hay calificaciones
- Describir Cada Uno de Los Pasos de Una Investigación de Brote.Documento12 páginasDescribir Cada Uno de Los Pasos de Una Investigación de Brote.Paul GarciaAún no hay calificaciones
- Caso 12Documento19 páginasCaso 12Paul GarciaAún no hay calificaciones
- Tratamientos Pulpares en OdontopediatríaDocumento13 páginasTratamientos Pulpares en OdontopediatríaPaul GarciaAún no hay calificaciones
- Componentes o Estructura de Un Artículo CientíficoDocumento2 páginasComponentes o Estructura de Un Artículo CientíficoPaul GarciaAún no hay calificaciones
- Acondicionadores de TejidosDocumento3 páginasAcondicionadores de TejidosPaul GarciaAún no hay calificaciones
- Material y MétodoDocumento1 páginaMaterial y MétodoPaul GarciaAún no hay calificaciones
- Uso de Farmacos en OdontopediatríaDocumento16 páginasUso de Farmacos en OdontopediatríaPaul GarciaAún no hay calificaciones
- Artículo Efectos de Provisionales-1Documento9 páginasArtículo Efectos de Provisionales-1Paul GarciaAún no hay calificaciones
- Acidosis Tubular Por Reflujo 2022Documento17 páginasAcidosis Tubular Por Reflujo 2022Paul GarciaAún no hay calificaciones
- Flora Bacteriana OralDocumento4 páginasFlora Bacteriana OralPaul GarciaAún no hay calificaciones
- Sondas PeriodontalesDocumento2 páginasSondas PeriodontalesPaul Garcia100% (1)
- Ligamento PeriodontalDocumento8 páginasLigamento PeriodontalPaul GarciaAún no hay calificaciones
- Universidad Nacional Autonoma de Nicaragua Unan-Leon: "A La Libertada Por La Universidad"Documento10 páginasUniversidad Nacional Autonoma de Nicaragua Unan-Leon: "A La Libertada Por La Universidad"Paul GarciaAún no hay calificaciones
- Enfermedades Metaxenicas Dengue OriginalDocumento31 páginasEnfermedades Metaxenicas Dengue OriginalKaren del Rocio Coronado YarlaqueAún no hay calificaciones
- Co Or063 07Documento10 páginasCo Or063 07Gabriel CadenaAún no hay calificaciones
- Reumatologia: Benemérita Universidad Autónoma de PueblaDocumento3 páginasReumatologia: Benemérita Universidad Autónoma de PueblaAiram ItzelAún no hay calificaciones
- Células Humanas Viejas Rejuvenecidas en Un Descubrimiento Revolucionario Sobre El EnvejecimientoDocumento27 páginasCélulas Humanas Viejas Rejuvenecidas en Un Descubrimiento Revolucionario Sobre El EnvejecimientoJean Romero YampulAún no hay calificaciones
- LeishmaniosisDocumento5 páginasLeishmaniosisAdrián Ramírez EsquivelAún no hay calificaciones
- El EosinofiloDocumento4 páginasEl Eosinofilopepe islasAún no hay calificaciones
- Antecedentes Clinicos Sobre LaserDocumento3 páginasAntecedentes Clinicos Sobre LaserCristhian Fernando Ñustes PeñaAún no hay calificaciones
- Apuntes Fisiopatologia I DR CordeiroDocumento41 páginasApuntes Fisiopatologia I DR CordeiroCAROLINA CALLIZAYA AVERANGAAún no hay calificaciones
- 3C - Paludismo 2020Documento12 páginas3C - Paludismo 2020Carlos Vinicio Chiluisa GuachoAún no hay calificaciones
- Sistema Linfatico e Inmunidad PDFDocumento134 páginasSistema Linfatico e Inmunidad PDFCarlos NietoAún no hay calificaciones
- Tema 5 - El Sistema InmuneDocumento14 páginasTema 5 - El Sistema Inmunepaco ruizAún no hay calificaciones
- Examen de InmunologiaDocumento10 páginasExamen de InmunologiaDante JosueAún no hay calificaciones
- TPM de Inmunología.Documento7 páginasTPM de Inmunología.Rut Ester González VásquezAún no hay calificaciones
- cuadernodedivulgacionRCP2015 2017 PDFDocumento78 páginascuadernodedivulgacionRCP2015 2017 PDFBoris TapiaAún no hay calificaciones
- Vademecum de MesoterapiaDocumento45 páginasVademecum de MesoterapiaRojas Evert Alonso67% (6)
- Inmunidad Frente A HongosDocumento1 páginaInmunidad Frente A HongosPaulina Sanchez SanchezAún no hay calificaciones
- TP12 HipersensibilidadDocumento19 páginasTP12 Hipersensibilidadmicro.fcvunlpAún no hay calificaciones
- Shock en Pediatria PDFDocumento84 páginasShock en Pediatria PDFalberto mendozaAún no hay calificaciones
- Aplasia MedularDocumento6 páginasAplasia MedularDavid Ross50% (2)
- PNI, David VinyesDocumento10 páginasPNI, David Vinyesquintero158Aún no hay calificaciones
- VIRBAC - INMUNOMODULADORES BovinoslecheDocumento8 páginasVIRBAC - INMUNOMODULADORES BovinoslecheRobinho RicardzAún no hay calificaciones
- Bonini Leonardi Los Aspectos Polifacéticos de Las Alergias Oculares - Fenotipos y Endotipos Conjuntivitis Alergica 2022.en - EsDocumento10 páginasBonini Leonardi Los Aspectos Polifacéticos de Las Alergias Oculares - Fenotipos y Endotipos Conjuntivitis Alergica 2022.en - EsCamila FlorezAún no hay calificaciones
- Cap.4 Trastornos Hemodinamicos, Tromboembolia y ShockDocumento86 páginasCap.4 Trastornos Hemodinamicos, Tromboembolia y ShockKevin Alejandro Arita ChinchillaAún no hay calificaciones
- Hypoderma MorrondoDocumento172 páginasHypoderma MorrondoJL LRAún no hay calificaciones
- Universidad Complutense de Madrid: Departamento de Nutrición, Bromatología y Tecnología de Los AlimentosDocumento247 páginasUniversidad Complutense de Madrid: Departamento de Nutrición, Bromatología y Tecnología de Los AlimentosIsabella RaquejoAún no hay calificaciones
- 9789591615978Documento148 páginas9789591615978Rafael Sauceda LopezAún no hay calificaciones
- Inmunidad Organoespecifica (Recuperado Automáticamente)Documento16 páginasInmunidad Organoespecifica (Recuperado Automáticamente)Valeria AmayaAún no hay calificaciones
- Metabolismo QuirúrgicoDocumento30 páginasMetabolismo QuirúrgicoCarlos ReyesAún no hay calificaciones
- Telma InmunólogiaDocumento356 páginasTelma InmunólogiaNicole AriasAún no hay calificaciones
- Reumato Final Ofi 2.0Documento233 páginasReumato Final Ofi 2.0Crixalis 145Aún no hay calificaciones