También podría gustarte
- Endotoxinas Usp 85Documento6 páginasEndotoxinas Usp 85orellanauvmcAún no hay calificaciones
- P2. Curva de Crecimiento BacterianoDocumento7 páginasP2. Curva de Crecimiento BacterianoJhohanAún no hay calificaciones
- Taller Indicadores Biológicos de PoluciónDocumento4 páginasTaller Indicadores Biológicos de PoluciónAndrea GarciaAún no hay calificaciones
- Manual Laboratorio de Microbiologia Industrial 2022Documento30 páginasManual Laboratorio de Microbiologia Industrial 2022isabell sanchez100% (1)
- Aplicación Muestreo.Documento2 páginasAplicación Muestreo.NICOLAS ACOSTA CUBILLOSAún no hay calificaciones
- Método 1682Documento20 páginasMétodo 1682Martin Mejia EscobarAún no hay calificaciones
- Cultivos de ProtoplastosDocumento35 páginasCultivos de ProtoplastosNormaAún no hay calificaciones
- Efecto Del Tiempo de Exposición y La Concentración de Un Tóxico Sobre El Desarrollo MicrobianoDocumento15 páginasEfecto Del Tiempo de Exposición y La Concentración de Un Tóxico Sobre El Desarrollo MicrobianoJENIFER PAOLA MENDOZA ISEDAAún no hay calificaciones
- Laboratorio 9 EcometraDocumento15 páginasLaboratorio 9 Ecometraarigatona0% (1)
- Tarea de Micro 2 (Mycoderma)Documento5 páginasTarea de Micro 2 (Mycoderma)maamago40Aún no hay calificaciones
- Agua PeptonadaDocumento2 páginasAgua PeptonadaalondraAún no hay calificaciones
- tesisUPV2468 VinosDocumento268 páginastesisUPV2468 VinosYrma Yenilda GmAún no hay calificaciones
- Tincion de Gram Informe TerminadoDocumento4 páginasTincion de Gram Informe TerminadoMiguel SantiagoAún no hay calificaciones
- Investigación Documental "Métodos Indirectos de Cuantificación de Microorganismos Viables y Totales"Documento5 páginasInvestigación Documental "Métodos Indirectos de Cuantificación de Microorganismos Viables y Totales"ArunAún no hay calificaciones
- Siembra de Aerobios Mesofilos Por El Metodo de Vertido en PlacaDocumento7 páginasSiembra de Aerobios Mesofilos Por El Metodo de Vertido en PlacaMiguel DrouetAún no hay calificaciones
- PDF Translator 1658442037311Documento2 páginasPDF Translator 1658442037311jimeAún no hay calificaciones
- La ZearalenonaDocumento4 páginasLa ZearalenonaDaniela1889Aún no hay calificaciones
- EmbDocumento3 páginasEmbana mariaAún no hay calificaciones
- PCR ANIDADA 1Documento3 páginasPCR ANIDADA 1Yôvanzhitô Cajo BarbozaAún no hay calificaciones
- Análisis Microbiológico de Turrones y MazapanesDocumento8 páginasAnálisis Microbiológico de Turrones y MazapanesannabethcamilaAún no hay calificaciones
- Análisis Microbiológico de Órganos de CamarónDocumento3 páginasAnálisis Microbiológico de Órganos de CamarónHilda FelixAún no hay calificaciones
- Cuantificacion de Polifenoles en Harina de PlatanoDocumento3 páginasCuantificacion de Polifenoles en Harina de PlatanoMeli CastañoAún no hay calificaciones
- Presentacion Caldo Rojo de FenolDocumento11 páginasPresentacion Caldo Rojo de FenolLuis Eduardo Delgado Ramirez50% (2)
- Informes HPLCDocumento6 páginasInformes HPLCMelani Herrera hernandezAún no hay calificaciones
- ntc-4779Documento30 páginasntc-4779Duban AvilaAún no hay calificaciones
- Parametros MicrobiologicosDocumento26 páginasParametros MicrobiologicosJorge Esteban RosalesAún no hay calificaciones
- NTC261 Aceite de ManiDocumento0 páginasNTC261 Aceite de ManiNicolas GranadosAún no hay calificaciones
- Yoshida 2016 Science Ideonella Sakaiensis Grows On PET - En.esDocumento5 páginasYoshida 2016 Science Ideonella Sakaiensis Grows On PET - En.esDeysi Yupanqui AroniAún no hay calificaciones
- RTD 334 Harinas Vegetales 1 Corregida 27 09 20111Documento14 páginasRTD 334 Harinas Vegetales 1 Corregida 27 09 20111JoseAún no hay calificaciones
- Anexo. Informe 2.obtencion de Enzimas y Tratamiento EnzimaticoDocumento16 páginasAnexo. Informe 2.obtencion de Enzimas y Tratamiento EnzimaticoDayana RealAún no hay calificaciones
- Cultivos IniciadoresDocumento24 páginasCultivos IniciadoresIsmael HernandezAún no hay calificaciones
- Laboratorio de Mohos y LevadurasDocumento3 páginasLaboratorio de Mohos y LevadurasIvan MoralesAún no hay calificaciones
- Apaza Choquehuanca Lourdes YasminDocumento120 páginasApaza Choquehuanca Lourdes YasminEnzo Sebastián AraujoAún no hay calificaciones
- Diagrama Estafilococos 0.1Documento1 páginaDiagrama Estafilococos 0.1Jennifer LinoAún no hay calificaciones
- Agar Salmonella ShigellaDocumento2 páginasAgar Salmonella ShigellaJorge CohenAún no hay calificaciones
- MicronúcleosDocumento6 páginasMicronúcleosDANIA CAROLINA GUTIERREZ HINOJOSA est100% (1)
- Ficha Técnica - Secador Por AsperciónDocumento3 páginasFicha Técnica - Secador Por AsperciónJesus Mesias IsraelAún no hay calificaciones
- NTC5478-pto HumoDocumento15 páginasNTC5478-pto HumoLuz Angela Diaz Guevara100% (2)
- Hemo BacterDocumento2 páginasHemo BacterLuz R. MendozaAún no hay calificaciones
- Recuento de Aerobios Mesofilos EsporuladosDocumento13 páginasRecuento de Aerobios Mesofilos EsporuladosLuis EgasAún no hay calificaciones
- Análisis Microbiológico de OperariosDocumento3 páginasAnálisis Microbiológico de OperariosNicol BuitragoAún no hay calificaciones
- Determinación de Coliformes TotalesDocumento8 páginasDeterminación de Coliformes TotalesPatrick PhoenixAún no hay calificaciones
- Mohos y Levaduras Esta de Aki Es La Ordenada Mejorada 100porcierto RealDocumento9 páginasMohos y Levaduras Esta de Aki Es La Ordenada Mejorada 100porcierto ReallolahAún no hay calificaciones
- Practica Staphylococcus AureusDocumento4 páginasPractica Staphylococcus Aureusdulzura123456789Aún no hay calificaciones
- Bioprocesos LevadurasDocumento4 páginasBioprocesos LevadurasAlejandro Guerra Rodriguez100% (1)
- Biorreactores en La Producción Del MezcalDocumento8 páginasBiorreactores en La Producción Del MezcalCésar Augusto JuárezAún no hay calificaciones
- Producción de Ácido Cítrico Por Fermentación en Estado Sólido Usando Bagazo de CañaDocumento4 páginasProducción de Ácido Cítrico Por Fermentación en Estado Sólido Usando Bagazo de CañaDiego Nicolas ManceraAún no hay calificaciones
- AISLAMIENTO DE Enterobacter Aerogenes FinalDocumento8 páginasAISLAMIENTO DE Enterobacter Aerogenes FinalTami MeyerAún no hay calificaciones
- Analisis Microbiologico de La LecheDocumento4 páginasAnalisis Microbiologico de La LecheMonica Patricia Rodriguez MachucaAún no hay calificaciones
- P2 Micro ExpDocumento4 páginasP2 Micro ExpClau VargasAún no hay calificaciones
- Nte Inen 1755 2 PDFDocumento15 páginasNte Inen 1755 2 PDFDiego VelezAún no hay calificaciones
- Justificación Del TrabajoDocumento1 páginaJustificación Del TrabajoAngel TapiaAún no hay calificaciones
- 2 Aerobios MesofilosDocumento5 páginas2 Aerobios MesofilosJennifer LinoAún no hay calificaciones
- Ocw Procesos Termicos ConservacionDocumento119 páginasOcw Procesos Termicos ConservacionJesus SandovalAún no hay calificaciones
- Práctica 3 - Aislamiento de LectinasDocumento2 páginasPráctica 3 - Aislamiento de LectinasDaniela IbarraAún no hay calificaciones
- Tema 8 TranscripcionDocumento15 páginasTema 8 Transcripcionjulian andresAún no hay calificaciones
- Marchitez de La PiñaDocumento14 páginasMarchitez de La PiñaBetssy Amparo Basurto RomeroAún no hay calificaciones
- Trabajo de GeneticaDocumento5 páginasTrabajo de GeneticaYucelis Paola Herazo SalgadoAún no hay calificaciones
- Virus RabiaDocumento36 páginasVirus RabiaMarcelo RealAún no hay calificaciones
- Semana 9 Clase 1 PB2Documento32 páginasSemana 9 Clase 1 PB2nayeli davilaAún no hay calificaciones
- Cristo No Pudo Haber Nacido El 25 de DiciembreDocumento33 páginasCristo No Pudo Haber Nacido El 25 de DiciembreDavid MendozaAún no hay calificaciones
- La Teoría de La Personalidad, Carl Rogers y George KellyDocumento14 páginasLa Teoría de La Personalidad, Carl Rogers y George KellyAdamary TobarAún no hay calificaciones
- Actividad Evaluable (A2) Sistema de Informacion Milagros GaleppiDocumento5 páginasActividad Evaluable (A2) Sistema de Informacion Milagros GaleppiRoxanaAún no hay calificaciones
- Fisica-Taller Prueba Saber 11°Documento14 páginasFisica-Taller Prueba Saber 11°Breyner Joel Ortiz DiazAún no hay calificaciones
- Bosquejo de Fin de AñoDocumento7 páginasBosquejo de Fin de AñoObed Pereyra100% (3)
- La Gastronomia en AsturiasDocumento15 páginasLa Gastronomia en AsturiasManolo RamosAún no hay calificaciones
- Tfg-N. 1498Documento79 páginasTfg-N. 1498Gerard Labernia TomasAún no hay calificaciones
- 22 Sesion de Aprendizaje Public Id AdDocumento7 páginas22 Sesion de Aprendizaje Public Id Adgabysantisteban6365100% (3)
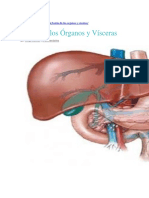
- Teoría Órganos y VíscerasDocumento6 páginasTeoría Órganos y VíscerasArq concAún no hay calificaciones
- Asertividad en Mi Vida Directiva, A RodriguezDocumento35 páginasAsertividad en Mi Vida Directiva, A RodriguezmarozzottoAún no hay calificaciones
- Clase 3 LecturaDocumento6 páginasClase 3 LecturaTania AcostaAún no hay calificaciones
- Accesorios de AudioDocumento3 páginasAccesorios de AudioArte Y MecatrónicaAún no hay calificaciones
- Reporte de BatelleDocumento3 páginasReporte de BatelleVianey GerardoAún no hay calificaciones
- Tecnicas de NegociacionDocumento1 páginaTecnicas de NegociacionAlejandro Garcìa0% (1)
- Report e Alum No Matric UlaDocumento1 páginaReport e Alum No Matric UlaBrayanAún no hay calificaciones
- Conocete A Ti MismoDocumento35 páginasConocete A Ti MismoIng Jimmy De la CruzAún no hay calificaciones
- MEC 36 - Pautas de Mantención Mecánica en Palas PHDocumento8 páginasMEC 36 - Pautas de Mantención Mecánica en Palas PHSebastian Alberto Tapia Rodriguez100% (1)
- Clasificaciones Con WekaDocumento15 páginasClasificaciones Con WekaveraluzmgAún no hay calificaciones
- Ensayo Proyecto de Cultura Nacional, Porfirio Diaz y La Modernizacion, Proyecto PosrevolucionarioDocumento7 páginasEnsayo Proyecto de Cultura Nacional, Porfirio Diaz y La Modernizacion, Proyecto PosrevolucionarioXinantecatl L SolaresAún no hay calificaciones
- Cuales Considera Que Son Los Problemas Ambientales Más Graves Que Afectan Nuestra ComunidadDocumento8 páginasCuales Considera Que Son Los Problemas Ambientales Más Graves Que Afectan Nuestra ComunidadElbys DelgadoAún no hay calificaciones
- Orden-Pre-252-2006 Modifica ITC APQ 10Documento4 páginasOrden-Pre-252-2006 Modifica ITC APQ 10Maria Fernanda ZuluagaAún no hay calificaciones
- Lorca MesinaDocumento147 páginasLorca Mesinamgarcia_244635Aún no hay calificaciones
- Manual Motor Diesel John Deere - Clarke PDFDocumento48 páginasManual Motor Diesel John Deere - Clarke PDFCésar Jc100% (2)
- Avances de Tesis UtrcDocumento12 páginasAvances de Tesis Utrcdavid arturo oriz rdzAún no hay calificaciones
- Presentac Planificac Control Gestión 31-10-22Documento16 páginasPresentac Planificac Control Gestión 31-10-22Arantxita OlivesAún no hay calificaciones
- Analisis de La Abeja HaraganaDocumento10 páginasAnalisis de La Abeja HaraganaSaray Beita JzAún no hay calificaciones
- Sistemas de Carburacion e InyeccionDocumento22 páginasSistemas de Carburacion e Inyeccionrafael perniaAún no hay calificaciones
- FOL06 TareaDocumento7 páginasFOL06 Tareaeduito1973100% (3)
- Trabajo de FutbolDocumento27 páginasTrabajo de FutbolMartha Elena0% (1)
- PlaneacionesDocumento22 páginasPlaneacionesEvelyn Parra100% (9)