También podría gustarte
- Crucigrama 2 AdultoDocumento1 páginaCrucigrama 2 AdultoEileen Caballero100% (1)
- Camara Frigorifica para Carne de CerdoDocumento36 páginasCamara Frigorifica para Carne de CerdoChristian SalasAún no hay calificaciones
- Motilidad GastrointestinalDocumento32 páginasMotilidad GastrointestinalJx JhoffanAún no hay calificaciones
- Nefropatia DiabeticaDocumento44 páginasNefropatia DiabeticaMatias VergaraAún no hay calificaciones
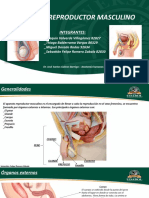
- Aparato Reproductor MasculinoDocumento23 páginasAparato Reproductor MasculinoDe todoo Un pocoAún no hay calificaciones
- GlicemiaDocumento84 páginasGlicemiaDante LuisAún no hay calificaciones
- Eje Hipotalamo - HipofisiarioDocumento28 páginasEje Hipotalamo - HipofisiarioVicky Padron100% (1)
- Malvern Teoria Difraccion EspañolDocumento9 páginasMalvern Teoria Difraccion EspañolMiguel Zuniga MarconiAún no hay calificaciones
- Minerales y Vitaminas en Rumiantes y No RumiantesDocumento28 páginasMinerales y Vitaminas en Rumiantes y No Rumiantesroberth paulAún no hay calificaciones
- Babesia EquinaDocumento35 páginasBabesia EquinaAlejandra Garzon RojasAún no hay calificaciones
- Endocrinologia Resumen Guyton e Hall Fisiologia Medica 13 EdDocumento25 páginasEndocrinologia Resumen Guyton e Hall Fisiologia Medica 13 EdRobert Mora TorosineAún no hay calificaciones
- Semana 12 Fisiología GastrointestinalDocumento78 páginasSemana 12 Fisiología GastrointestinalAline AyoneAún no hay calificaciones
- Sistema Hematopoyético GoodDocumento181 páginasSistema Hematopoyético GoodAmalia VillavicencioAún no hay calificaciones
- Embriologia Del Sistema RespiratorioDocumento20 páginasEmbriologia Del Sistema RespiratorioAlessia BeckerAún no hay calificaciones
- Anatomia y Fisiologia Del Aparato DigestivoDocumento13 páginasAnatomia y Fisiologia Del Aparato DigestivoAndrea Rojas SeminarioAún no hay calificaciones
- Aparato Reproductor Del MachoDocumento45 páginasAparato Reproductor Del MachoBastianAún no hay calificaciones
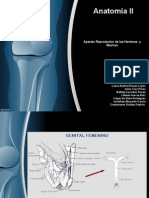
- Genital Bovino Masculino Anatomia IIDocumento119 páginasGenital Bovino Masculino Anatomia IIriodulce117Aún no hay calificaciones
- Resumen Metazoos IntestinalesDocumento17 páginasResumen Metazoos IntestinalesAngelik bonillaAún no hay calificaciones
- Repaso-Patología Sistémica VeterinariaDocumento24 páginasRepaso-Patología Sistémica Veterinarialuis javierAún no hay calificaciones
- 6 Pancreas EndocrinoDocumento53 páginas6 Pancreas EndocrinoFabiola ChadidAún no hay calificaciones
- Anatomía y Fisiología de Los RiñonesDocumento18 páginasAnatomía y Fisiología de Los RiñonesDante LuisAún no hay calificaciones
- Incrustaciones (INLAYS, OnLAYS)Documento14 páginasIncrustaciones (INLAYS, OnLAYS)Maileth Nasner Quintero100% (5)
- ProteinasDocumento6 páginasProteinasRodrigo LopezAún no hay calificaciones
- Aparato Reproductor Hembra y MachoDocumento72 páginasAparato Reproductor Hembra y MachoNallely GlezAún no hay calificaciones
- Tema 28 Fisiologia de Las Hormonas FemeninasDocumento12 páginasTema 28 Fisiologia de Las Hormonas FemeninasElisinaAún no hay calificaciones
- Glándulas y SecreciónDocumento26 páginasGlándulas y SecreciónMariana VillarrealAún no hay calificaciones
- Histologia Sistema Endocrino Acb PDFDocumento78 páginasHistologia Sistema Endocrino Acb PDFWildercino RAAún no hay calificaciones
- Clase8 Pisces VillenaArboccóDocumento30 páginasClase8 Pisces VillenaArboccóJhannaAún no hay calificaciones
- Anatomia Del CorazónDocumento1 páginaAnatomia Del CorazónNathalie DávilaAún no hay calificaciones
- Resumen Fisiología Del MúsculoDocumento10 páginasResumen Fisiología Del MúsculoErika Sanchez H0% (1)
- Marco TeóricoDocumento24 páginasMarco TeóricoJohan AndradeAún no hay calificaciones
- Anatomia de La GallinaDocumento28 páginasAnatomia de La Gallinavictor alpirezAún no hay calificaciones
- PlacentacionDocumento23 páginasPlacentacionHugo Quiles DetweilerAún no hay calificaciones
- Quimica SanguineaDocumento10 páginasQuimica SanguineaLuis RiveraAún no hay calificaciones
- Histología Del OídoDocumento11 páginasHistología Del OídoluisAún no hay calificaciones
- HEMOSTASIADocumento15 páginasHEMOSTASIAJhonn Paolo Flores Barreto Xelnax100% (1)
- Ojo Anatomía e Histología, FinalDocumento6 páginasOjo Anatomía e Histología, FinalJacob MendozaAún no hay calificaciones
- Sistema CirculatorioDocumento104 páginasSistema CirculatorioBerenice Hernández EnríquezAún no hay calificaciones
- Insuficiencia CardiacaDocumento34 páginasInsuficiencia CardiacamedinahhAún no hay calificaciones
- Apuntes de Farmacologia MayraDocumento39 páginasApuntes de Farmacologia MayraErika R. Zazueta LéonAún no hay calificaciones
- Cuadro Comparativo EndocrinoDocumento7 páginasCuadro Comparativo EndocrinoTanya Rubí100% (1)
- Actividad Osmotica Del RiñonDocumento10 páginasActividad Osmotica Del RiñonMiguel Bolsa PeñaAún no hay calificaciones
- Produccion PorcinaDocumento12 páginasProduccion PorcinaJorge Luis Ramirez BenitezAún no hay calificaciones
- Rickettsias 120510220539 Phpapp01Documento27 páginasRickettsias 120510220539 Phpapp01H3inzzAún no hay calificaciones
- Intestino DelgadoDocumento22 páginasIntestino DelgadoIvette Avendaño MejíaAún no hay calificaciones
- El Tubo Intestinal y Las Cavidades CorporalesDocumento48 páginasEl Tubo Intestinal y Las Cavidades CorporalesUlises García Arellano100% (7)
- Fisiologia GastrointestinalDocumento2 páginasFisiologia Gastrointestinaljosep_djmc9424Aún no hay calificaciones
- Facultad de Medicina Veterinaria Y Zootecnia: "Sistema NerviosoDocumento29 páginasFacultad de Medicina Veterinaria Y Zootecnia: "Sistema NerviosoLeonel Villaseñor CorteroAún no hay calificaciones
- Función Tubular UNEFM Pedro RodríguezDocumento8 páginasFunción Tubular UNEFM Pedro RodríguezPedro RodriguezAún no hay calificaciones
- Las Funciones de Los Diverticulos Del RumiantesDocumento6 páginasLas Funciones de Los Diverticulos Del Rumiantesedgar cundapiAún no hay calificaciones
- CLASE 7. Hematopoyesis. Funciones, Características Físicas de La Sangre. Constitución Anatómica.Documento26 páginasCLASE 7. Hematopoyesis. Funciones, Características Físicas de La Sangre. Constitución Anatómica.Juan SandovalAún no hay calificaciones
- Hidatidosis 2016Documento55 páginasHidatidosis 2016julioAún no hay calificaciones
- Tubo DigestivoDocumento229 páginasTubo DigestivoRita Alexandra Murillo VillamarAún no hay calificaciones
- Fisiologia Del VomitoDocumento3 páginasFisiologia Del VomitoAlexandra Olortegui MendietaAún no hay calificaciones
- GENERALIDADES DE OSTEOLOGÍA 2020 - 2 Guia 2Documento7 páginasGENERALIDADES DE OSTEOLOGÍA 2020 - 2 Guia 2Laura Sofia Avendaño MoralesAún no hay calificaciones
- Glándulas SalivalesDocumento46 páginasGlándulas SalivalesJessica0% (1)
- CéstodosDocumento28 páginasCéstodosDaniela VinuezaAún no hay calificaciones
- 2 Unidad2 Tejido GlandularDocumento10 páginas2 Unidad2 Tejido Glandularluna2702Aún no hay calificaciones
- Hembra de Shivelly AnatomiaDocumento10 páginasHembra de Shivelly AnatomiaLuis Miguel AmayaAún no hay calificaciones
- CascoDocumento14 páginasCascoAgus DíazAún no hay calificaciones
- Tumor Venerio Transmisible TVT PerrosDocumento16 páginasTumor Venerio Transmisible TVT PerrosluisAún no hay calificaciones
- A9 MFNLDocumento4 páginasA9 MFNLmaria fernanda najera lunaAún no hay calificaciones
- Fluidoterapia en Animales MenoresDocumento62 páginasFluidoterapia en Animales MenoresSandra Montes FaustorAún no hay calificaciones
- Guía Enfermedades Infecciosas 2013-IIDocumento19 páginasGuía Enfermedades Infecciosas 2013-IIAlexx RomarAún no hay calificaciones
- Renal HidrosalinoDocumento37 páginasRenal HidrosalinoViviana Hernandez QuijanoAún no hay calificaciones
- Aparato UrinarioDocumento5 páginasAparato UrinarioDaniel MéndezAún no hay calificaciones
- Clase 1 y 2Documento43 páginasClase 1 y 2studyshop29Aún no hay calificaciones
- HEMOCULTIVODocumento9 páginasHEMOCULTIVODante LuisAún no hay calificaciones
- Creatin in ADocumento11 páginasCreatin in ADante LuisAún no hay calificaciones
- Histologia Del RiñonDocumento4 páginasHistologia Del RiñonDante LuisAún no hay calificaciones
- Concreto Vaciado en SitioDocumento13 páginasConcreto Vaciado en SitioHeriberto Yau BAún no hay calificaciones
- Tema 1 Introducción A La RMDocumento83 páginasTema 1 Introducción A La RMManuelAún no hay calificaciones
- Guía Nº01 Cemento.Documento12 páginasGuía Nº01 Cemento.Digna VilchezAún no hay calificaciones
- Conceptos Básicos de Toxicología LaboralDocumento8 páginasConceptos Básicos de Toxicología LaboralSebastian Andres Gonzalez ValdiviaAún no hay calificaciones
- Práctica Filtración Arena y Torta 2023-24Documento9 páginasPráctica Filtración Arena y Torta 2023-24dayana.remigio96Aún no hay calificaciones
- Duraplate Es 301 SwaDocumento4 páginasDuraplate Es 301 SwaGiancarlo Sanchez EspinozaAún no hay calificaciones
- 04 PH NALCO OKDocumento10 páginas04 PH NALCO OKDavid Cruz ZamoraAún no hay calificaciones
- Grasso SPduoDocumento90 páginasGrasso SPduoEduardo Manuel Leite LopesAún no hay calificaciones
- Anteproyecto Tesis Efectividad de La Deshidratacion OsmoticaDocumento21 páginasAnteproyecto Tesis Efectividad de La Deshidratacion OsmoticaJorge Brian Zapata BoyerAún no hay calificaciones
- Síntesis de Bromuro de TercDocumento5 páginasSíntesis de Bromuro de TercAlejandra De la CruzAún no hay calificaciones
- ESTRATEGIAS DE ALIMENTACIÓN DE GANADO DE Carne de Feetlot PDFDocumento3 páginasESTRATEGIAS DE ALIMENTACIÓN DE GANADO DE Carne de Feetlot PDFFercho Lotudo100% (1)
- Hoja de Datos Del Tic106Documento9 páginasHoja de Datos Del Tic106Jesus Andres Intriago VelezAún no hay calificaciones
- El Bocadillo de Guayaba Se Elabora en Varios Países de América LatinaDocumento3 páginasEl Bocadillo de Guayaba Se Elabora en Varios Países de América LatinaJaneth PeñaAún no hay calificaciones
- PRACTICA N 03 y 04Documento13 páginasPRACTICA N 03 y 04Yhán AnthonyAún no hay calificaciones
- Análisis VolumétricoDocumento33 páginasAnálisis VolumétricoToddCrahanAún no hay calificaciones
- Informe de Salto HidraulicoDocumento11 páginasInforme de Salto HidraulicoEnrique VigilAún no hay calificaciones
- Marco Teórico PectinaDocumento28 páginasMarco Teórico PectinasernajaimeAún no hay calificaciones
- Agrotecnia - Labranza Del SueloDocumento9 páginasAgrotecnia - Labranza Del SueloJose HenrryAún no hay calificaciones
- Compendio Qui 125 2º Prueba Catedra 2007Documento36 páginasCompendio Qui 125 2º Prueba Catedra 2007Adriana Toro RosalesAún no hay calificaciones
- Presentacion Sobre Tesis Sobre El CilantronDocumento45 páginasPresentacion Sobre Tesis Sobre El Cilantronni2bthAún no hay calificaciones
- DEFINICIONESDocumento4 páginasDEFINICIONESMonito OrdoñezAún no hay calificaciones
- Resumen de CombustiblesDocumento3 páginasResumen de CombustiblesManuel Herrera100% (1)
- U4 Hidrostatica 2018 PDFDocumento4 páginasU4 Hidrostatica 2018 PDFgonzaloch8Aún no hay calificaciones
- Tungsteno Hoja de SeguridadDocumento19 páginasTungsteno Hoja de SeguridadAlma SánchezAún no hay calificaciones
- NutracéuticosDocumento4 páginasNutracéuticossusanamapazAún no hay calificaciones