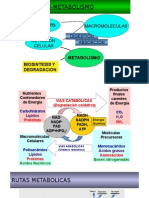
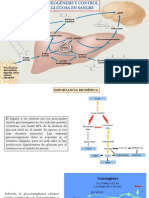
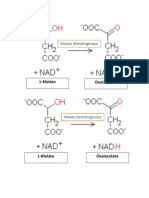
También podría gustarte
- Ciclo de La PentosaDocumento46 páginasCiclo de La PentosaLuis Contreras ArtuzAún no hay calificaciones
- GlucolisisDocumento15 páginasGlucolisisalejo_1994244644100% (1)
- Nutrición y rutas metabólicas claveDocumento56 páginasNutrición y rutas metabólicas claveXavi EscobarAún no hay calificaciones
- GlucólisisDocumento52 páginasGlucólisisEduardo Limay Coronado100% (4)
- Transcripción y TraducciónDocumento8 páginasTranscripción y TraducciónMariana Restrepo Benavides0% (1)
- Estructura Membrana CelularDocumento3 páginasEstructura Membrana CelularALFREDOAún no hay calificaciones
- Clase de Las Pentosa FosfatoDocumento18 páginasClase de Las Pentosa FosfatoJesus RamirezAún no hay calificaciones
- GlucolisisDocumento26 páginasGlucolisisAle JoyaAún no hay calificaciones
- Glucólisis: 10 reacciones para convertir glucosa en piruvato generando ATPDocumento1 páginaGlucólisis: 10 reacciones para convertir glucosa en piruvato generando ATPDavid100% (1)
- Pentosas FosfatoDocumento39 páginasPentosas FosfatoKarolAún no hay calificaciones
- Glucogenética: Síntesis y degradación del glucógenoDocumento2 páginasGlucogenética: Síntesis y degradación del glucógenoGexAún no hay calificaciones
- BIOQUIMICA Ciclo de KrebsDocumento74 páginasBIOQUIMICA Ciclo de KrebsleonelAún no hay calificaciones
- Pentosas FosfatoDocumento21 páginasPentosas FosfatoSlovan Ochoa100% (1)
- Biosíntesis de Purinas y PirimidinasDocumento58 páginasBiosíntesis de Purinas y PirimidinasAnthony ARAún no hay calificaciones
- GLUCONEOGENESISDocumento32 páginasGLUCONEOGENESISzatanna66100% (2)
- Ciclo de La Pentosa FosfatoDocumento7 páginasCiclo de La Pentosa FosfatoeiderAún no hay calificaciones
- B-Oxidacion y Sintesis de Acidos GrasosDocumento22 páginasB-Oxidacion y Sintesis de Acidos GrasosPatricio Coronado75% (8)
- Beta oxidación de ácidos grasos: proceso catabólico por ciclos de 4 reaccionesDocumento16 páginasBeta oxidación de ácidos grasos: proceso catabólico por ciclos de 4 reaccionesshirley100% (1)
- Ciclo de La UreaDocumento6 páginasCiclo de La UreaChristian GavilánezAún no hay calificaciones
- Tema 13 Ruta Pentosa-FosfatoDocumento6 páginasTema 13 Ruta Pentosa-FosfatoIsismar VillamizarAún no hay calificaciones
- Vias de Las PentosasDocumento36 páginasVias de Las PentosasChristian Flores Beltrán100% (1)
- Glucogenolisis Glucogénesis Cetogénesis 5Documento45 páginasGlucogenolisis Glucogénesis Cetogénesis 5DianaOrdoñez100% (1)
- GLUCÓLISISDocumento15 páginasGLUCÓLISISDylan EstebanAún no hay calificaciones
- Vía pentosa fosfatoDocumento18 páginasVía pentosa fosfatoAndré HerreraAún no hay calificaciones
- La glucolisis: 10 pasos para generar energíaDocumento18 páginasLa glucolisis: 10 pasos para generar energíaOscar Antunez BitesAún no hay calificaciones
- Ruta de Las Pentosas FosfatoDocumento21 páginasRuta de Las Pentosas FosfatoecheegarayAún no hay calificaciones
- Ciclo de La UreaDocumento4 páginasCiclo de La UreaDaniel Salazar100% (1)
- La Ruta de Las Pentosas FosfatoDocumento4 páginasLa Ruta de Las Pentosas FosfatoMiguel MuñozAún no hay calificaciones
- Via de Las PentosasDocumento24 páginasVia de Las PentosasRoy Michael Guerra ArandaAún no hay calificaciones
- Ruta Pentosa-Fosfato: Producción de NADPH y RibosaDocumento5 páginasRuta Pentosa-Fosfato: Producción de NADPH y RibosaMarilin Blanco100% (1)
- Vía de Las PentosasDocumento36 páginasVía de Las PentosasLuis Rodolfo LópezAún no hay calificaciones
- Purinas y PirimidinasDocumento15 páginasPurinas y PirimidinasCristian Fernando OrtizAún no hay calificaciones
- Glucólisis, Gluconeogénesis y Ruta de Las Pentosas FosfatoDocumento26 páginasGlucólisis, Gluconeogénesis y Ruta de Las Pentosas FosfatoMarcelo Kim100% (1)
- Ruta de Las Pentosas FosfatoDocumento10 páginasRuta de Las Pentosas FosfatoAlexiiss LuiissAún no hay calificaciones
- Gluconeogenesis y Control de La Glucosa en SangreDocumento37 páginasGluconeogenesis y Control de La Glucosa en SangreStefano GarciaAún no hay calificaciones
- Vía de Las PentosasDocumento30 páginasVía de Las PentosasAlexia Fuentes100% (1)
- Via de Las Pentosas Fosfato y Nadph (Harvey Unidad 13)Documento8 páginasVia de Las Pentosas Fosfato y Nadph (Harvey Unidad 13)IlderAlvaradoAún no hay calificaciones
- Lípidos esenciales enDocumento51 páginasLípidos esenciales enDaniel LunaAún no hay calificaciones
- Regulación Hormonal de CarbohidratosDocumento5 páginasRegulación Hormonal de CarbohidratosDaniel RutledgeAún no hay calificaciones
- Vía de Las Pentosas FosfatoDocumento12 páginasVía de Las Pentosas FosfatoJosé AlfredoAún no hay calificaciones
- Metabolismo Del Glucogeno (Presentación)Documento27 páginasMetabolismo Del Glucogeno (Presentación)Guadalupe RiveraAún no hay calificaciones
- Vía de Las Pentosas FosfatoDocumento8 páginasVía de Las Pentosas Fosfatobrenda reynosoAún no hay calificaciones
- Fosforilación OxidativaDocumento8 páginasFosforilación OxidativaLourdes PerdomoAún no hay calificaciones
- GLUCONEOGENESISDocumento4 páginasGLUCONEOGENESISmiccxoAún no hay calificaciones
- La Enzima Malato DeshidrogenasaDocumento2 páginasLa Enzima Malato DeshidrogenasaKaren Torres JuárezAún no hay calificaciones
- GlucólisisDocumento26 páginasGlucólisislautarok100% (2)
- Metabolismo de NucleótidosDocumento8 páginasMetabolismo de NucleótidosAnonymous woR2GSAún no hay calificaciones
- Ciclo de La GlucolisisDocumento13 páginasCiclo de La GlucolisisCriss AdrianAún no hay calificaciones
- Reacciones de AminoazúcaresDocumento2 páginasReacciones de AminoazúcaresAbner RodriguezAún no hay calificaciones
- GLUCOGENOLISISDocumento6 páginasGLUCOGENOLISISCristobal Gamboa CamposAún no hay calificaciones
- La Ruta de Las Pentosas FosfatoDocumento47 páginasLa Ruta de Las Pentosas FosfatoSergio Emilio Fllres FuentesAún no hay calificaciones
- Via de La GluconeogenesisDocumento38 páginasVia de La Gluconeogenesisfmh200967% (6)
- Vía de Las Pentosas FosfatoDocumento9 páginasVía de Las Pentosas FosfatoJeff Willian Rosas QuinaAún no hay calificaciones
- Sintesis de TrigliceridosDocumento5 páginasSintesis de Trigliceridoskatiyerar100% (3)
- Metabolismo lípidos membranasDocumento3 páginasMetabolismo lípidos membranasErnesto MenendezAún no hay calificaciones
- Bioquimica Via de Pentosas FosfatoDocumento21 páginasBioquimica Via de Pentosas FosfatoLucero LópezAún no hay calificaciones
- Glucolisis 21645 PDFDocumento77 páginasGlucolisis 21645 PDFShärlÿ ContrerasAún no hay calificaciones
- El ciclo de la urea: transformación del amoníaco en ureaDocumento65 páginasEl ciclo de la urea: transformación del amoníaco en ureaAlexandraMedinaCastilloAún no hay calificaciones
- Aminoacidos No Esenciales (Expo)Documento16 páginasAminoacidos No Esenciales (Expo)AlbaOsorioAún no hay calificaciones
- Metabolismo de EicosanoidesDocumento16 páginasMetabolismo de EicosanoidesGino Chávez SánchezAún no hay calificaciones
- FOSFORILACIÓN OXIDATIVA Y TRANSPORTE DE ELECTRONESDocumento32 páginasFOSFORILACIÓN OXIDATIVA Y TRANSPORTE DE ELECTRONESUriel AlvaradoAún no hay calificaciones
- Tp9. Glucidos 2Documento18 páginasTp9. Glucidos 2ferminsAún no hay calificaciones
- Reforzamiento Academico Sem 04Documento4 páginasReforzamiento Academico Sem 04Ander PacoriAún no hay calificaciones
- Bioquímica muscular post-mortemDocumento31 páginasBioquímica muscular post-mortemKarol ObandoAún no hay calificaciones
- Histologia - 1er Parcial - Simulacro Preparcial AlumedDocumento40 páginasHistologia - 1er Parcial - Simulacro Preparcial AlumedAsi AsaAún no hay calificaciones
- Taller Biologia PDFDocumento3 páginasTaller Biologia PDFWalton CordobaAún no hay calificaciones
- Herencia MaternaDocumento4 páginasHerencia MaternaLuis CafAún no hay calificaciones
- Las ChaperoninasDocumento7 páginasLas ChaperoninasGerson Tocno LopezAún no hay calificaciones
- AtpDocumento18 páginasAtpLuis CorderoAún no hay calificaciones
- Resumen Final Biol. Cel. y Molec.Documento211 páginasResumen Final Biol. Cel. y Molec.Hugo MedusAún no hay calificaciones
- Horarios Quinto Semestre Septiembre 2108-Mayo 2019.Documento2 páginasHorarios Quinto Semestre Septiembre 2108-Mayo 2019.Daniel CisnerosAún no hay calificaciones
- Examen preparatorio UnivalleDocumento42 páginasExamen preparatorio UnivalleMaximiliano Cortes Tapia100% (1)
- Cuestionario Fisiologia Del Sistema Vascular Unidad 1Documento13 páginasCuestionario Fisiologia Del Sistema Vascular Unidad 1Arely CierraAún no hay calificaciones
- Funciones Biológicas de Las Proteínas - BiologuiasDocumento6 páginasFunciones Biológicas de Las Proteínas - BiologuiasMaryuris HernandezAún no hay calificaciones
- Prueba Quinto Basico CelulaDocumento4 páginasPrueba Quinto Basico Celulamassielfuentes_cedelAún no hay calificaciones
- Sistemas de defensa del organismoDocumento32 páginasSistemas de defensa del organismoweikeAún no hay calificaciones
- UntitledDocumento646 páginasUntitledNatalia Reifschneider100% (1)
- Banco de Preguntas LoliDocumento38 páginasBanco de Preguntas Loligaby_diaz_739Aún no hay calificaciones
- Biomoléculas orgánicas: carbohidratos, proteínas, lípidos y ácidos nucleicosDocumento6 páginasBiomoléculas orgánicas: carbohidratos, proteínas, lípidos y ácidos nucleicosPaulina MatamorosAún no hay calificaciones
- Bioelementos - Celula11Documento55 páginasBioelementos - Celula11Jhoselyn AlmeidaAún no hay calificaciones
- Digestión y Oxidación LípidosDocumento35 páginasDigestión y Oxidación LípidosasasaAún no hay calificaciones
- Hla y EmbarazoDocumento11 páginasHla y EmbarazoLudmir max Pascual roblesAún no hay calificaciones
- Metabolismo de Lipidos 1.1 1Documento46 páginasMetabolismo de Lipidos 1.1 1Sheyla Espino RamosAún no hay calificaciones
- Actividad 1. n1. Ciclo Vi BioDocumento3 páginasActividad 1. n1. Ciclo Vi BiotatianmacaAún no hay calificaciones
- Respiracion CelularDocumento7 páginasRespiracion CelularCristian VillarragaAún no hay calificaciones
- Guía estudio primer parcial nutriciónDocumento2 páginasGuía estudio primer parcial nutriciónLaura AcostaAún no hay calificaciones
- Musculos FuncionDocumento23 páginasMusculos Funciondilenia guzmanAún no hay calificaciones
- Bioquimica 3Documento14 páginasBioquimica 3Jorge RibonAún no hay calificaciones
- P1 BiologíaDocumento16 páginasP1 BiologíaPUNTAJE NACIONALAún no hay calificaciones
- Evaluación 1 - Transporte de sustancias a través de membranasDocumento9 páginasEvaluación 1 - Transporte de sustancias a través de membranasPaula ValdèsAún no hay calificaciones