También podría gustarte
- Anatomía Patológica de EstafilococosDocumento21 páginasAnatomía Patológica de EstafilococosNichole BrownAún no hay calificaciones
- Copia de Corazón Aislado de Sapo y Corazon in Situ StanniusDocumento3 páginasCopia de Corazón Aislado de Sapo y Corazon in Situ StanniusAlexis Moreno0% (1)
- Desórdenes en Receptores CelularesDocumento40 páginasDesórdenes en Receptores CelularesMarco MadridAún no hay calificaciones
- Glosario FisiopatologíaDocumento4 páginasGlosario FisiopatologíamassielvillalobosAún no hay calificaciones
- Alteraciones SubcelularesDocumento32 páginasAlteraciones SubcelularesFernanda Nicole Castillo Parra67% (3)
- Vena Cava Superior DobleDocumento3 páginasVena Cava Superior Doblejuliana pabon50% (2)
- DGB y DGT PresentaciónDocumento36 páginasDGB y DGT PresentaciónEdgaris Sc'r0% (1)
- Cuestionario de Imagenologia (Abdomen)Documento3 páginasCuestionario de Imagenologia (Abdomen)Krishna Leiva SilvaAún no hay calificaciones
- Aislamiento y Cultivo de Linfocitos y Neutrofilos HumanosDocumento5 páginasAislamiento y Cultivo de Linfocitos y Neutrofilos HumanoscrisleelopezAún no hay calificaciones
- Fisiologia Regulaciondelarespiracion 140916092221 Phpapp01Documento2 páginasFisiologia Regulaciondelarespiracion 140916092221 Phpapp01Fabiola SanchezAún no hay calificaciones
- Balotario Fisiologia FinalDocumento14 páginasBalotario Fisiologia FinalArlet Kristel Remuzgo Alvarado100% (1)
- Estudio de Caso HidrocefaliaDocumento17 páginasEstudio de Caso HidrocefaliaYenni Marcela TORRES NUNEZAún no hay calificaciones
- MAMA y CordonDocumento5 páginasMAMA y CordonDaniela PerezAún no hay calificaciones
- Embriologia de ToraxDocumento4 páginasEmbriologia de Toraxapi-19734073Aún no hay calificaciones
- Bioquimica de La Caries Final 2018Documento12 páginasBioquimica de La Caries Final 2018Diego Castaño MejiaAún no hay calificaciones
- Los Aminoácidos y Las Proteínas Como Electrolitos y Su Capacidad Amortiguadora en El Organismo HumanoDocumento12 páginasLos Aminoácidos y Las Proteínas Como Electrolitos y Su Capacidad Amortiguadora en El Organismo HumanoMiguel Ángel Jesús Lavarello BermejoAún no hay calificaciones
- 2 Proceso Salud y Enfermedad Determinantes de La SaludDocumento17 páginas2 Proceso Salud y Enfermedad Determinantes de La SaludNorma GuadalupeAún no hay calificaciones
- Sistema Del Complemento: Vías de ActivaciónDocumento6 páginasSistema Del Complemento: Vías de ActivaciónDaniela PonceAún no hay calificaciones
- Inflamación y Reparación TisularDocumento11 páginasInflamación y Reparación TisularBetty CarreñoAún no hay calificaciones
- Reporte de Bioquimica 8Documento4 páginasReporte de Bioquimica 8Lalo CarvajalAún no hay calificaciones
- Virus Lentos No ConvencionalesDocumento11 páginasVirus Lentos No Convencionalesmiguel110198250% (2)
- MONOGRAFÍA El Sentido Del OlfatoDocumento24 páginasMONOGRAFÍA El Sentido Del OlfatoOlga Aguirre RedondoAún no hay calificaciones
- Capítulo 46Documento26 páginasCapítulo 46Israel Arroyo Velazquez100% (4)
- Banco de PreguntasDocumento3 páginasBanco de PreguntasMarcelaAún no hay calificaciones
- MONOGRAFIA Esclerosis Lateral 2Documento13 páginasMONOGRAFIA Esclerosis Lateral 2Suzette XZ0% (1)
- Informe Fisiología Digestivo IIDocumento12 páginasInforme Fisiología Digestivo IIAna Belén Aguilar Pilco0% (1)
- OSMOSIS Y BASES BIOLOGICAS para El AlumnoDocumento30 páginasOSMOSIS Y BASES BIOLOGICAS para El AlumnoMarianela Rios Aillon100% (1)
- Grupo 11 Tejido Hematopoyético PreguntasDocumento2 páginasGrupo 11 Tejido Hematopoyético PreguntasMateoo06Aún no hay calificaciones
- Cap 76 Hormona Hipofisarias y Su Control Por El HipotálamoDocumento24 páginasCap 76 Hormona Hipofisarias y Su Control Por El Hipotálamofisiol100% (1)
- Termometria ClinicaDocumento8 páginasTermometria ClinicaJJJonathanAún no hay calificaciones
- Sílabo Bases Moleculares y Celulares de La Medicina I 2023-IDocumento19 páginasSílabo Bases Moleculares y Celulares de La Medicina I 2023-IDaniela Paredes gonzales100% (1)
- Inmunoglobulinas y AnticuerposDocumento20 páginasInmunoglobulinas y Anticuerposmichel100% (1)
- Metabolismo CerebralDocumento12 páginasMetabolismo CerebralFranklinYanes100% (3)
- Marco Teorico y Antecedentes-EthelDocumento22 páginasMarco Teorico y Antecedentes-EtheljeancAún no hay calificaciones
- Glosario de Términos Utilizados en Biología CelularDocumento3 páginasGlosario de Términos Utilizados en Biología CelularAlejandro López PedrazaAún no hay calificaciones
- Habito Corporal PDFDocumento8 páginasHabito Corporal PDFAnonymous grRZ9X45kAún no hay calificaciones
- Tecnicas As - Reacciones 2dariasDocumento4 páginasTecnicas As - Reacciones 2dariasdarkzero_858Aún no hay calificaciones
- Shock EspinalDocumento17 páginasShock EspinalAnalitha KnivesAún no hay calificaciones
- Menciones Las Características Generales Del Tejido EpitelialDocumento6 páginasMenciones Las Características Generales Del Tejido EpitelialOscar VasquezAún no hay calificaciones
- Segundos MensajerosDocumento14 páginasSegundos MensajerosArnold Gomez AlvaAún no hay calificaciones
- Síndrome de Cráneo en TrébolDocumento13 páginasSíndrome de Cráneo en TrébolTANIA JHANET CONDORI CONDORIAún no hay calificaciones
- Shock EspinalDocumento36 páginasShock EspinalFernando Antonio YarascaAún no hay calificaciones
- Practica 3 - Patologia Practica - Dr. RacchumiDocumento14 páginasPractica 3 - Patologia Practica - Dr. RacchumiAngie Deza GuzmanAún no hay calificaciones
- Examen de FisiopatologiaDocumento6 páginasExamen de FisiopatologiaHugo Quispe100% (1)
- EosinofilosDocumento4 páginasEosinofilosEstefy JerezAún no hay calificaciones
- Capitulo 20 FisiologiaDocumento30 páginasCapitulo 20 FisiologiaBrenda Mota100% (1)
- Anemias HarrisonDocumento9 páginasAnemias HarrisonAndrea GallegoAún no hay calificaciones
- Práctica #1 de Fisiología Neuromuscular y de Los Sistemas SensorialesDocumento6 páginasPráctica #1 de Fisiología Neuromuscular y de Los Sistemas SensorialesANDREA ZENA IMANAún no hay calificaciones
- Factores de Crecimiento HematopoyeticosDocumento1 páginaFactores de Crecimiento Hematopoyeticossisapuchaicela100% (3)
- M InferiorDocumento16 páginasM InferiorNora Maricela Cavazos MendozaAún no hay calificaciones
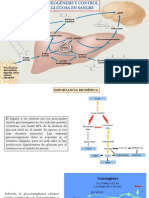
- Gluconeogenesis y Control de La Glucosa en SangreDocumento37 páginasGluconeogenesis y Control de La Glucosa en SangreStefano GarciaAún no hay calificaciones
- My Co BacteriasDocumento36 páginasMy Co BacteriasChateo Con QuienceaAún no hay calificaciones
- Informe Caso 1Documento12 páginasInforme Caso 1Valeria Tovar MartinezAún no hay calificaciones
- Practica 04 Métodos EstadísticosDocumento3 páginasPractica 04 Métodos EstadísticosAntonyAún no hay calificaciones
- Autoevaluacion Huesos Del CraneoDocumento4 páginasAutoevaluacion Huesos Del CraneoMarco Gonzales SilvaAún no hay calificaciones
- Historia de La Prevención de EnfermedadesDocumento26 páginasHistoria de La Prevención de EnfermedadesDani CicardiniAún no hay calificaciones
- 1.02 Respiracion CelularDocumento10 páginas1.02 Respiracion CelularAnonymous K8fT6HAún no hay calificaciones
- Cadena de Transporte de ElectronesDocumento10 páginasCadena de Transporte de ElectronesBelem ValenciaAún no hay calificaciones
- Fosforilación OxidativaDocumento25 páginasFosforilación OxidativaEmilio Jeanpaul Mitacc GutierrezAún no hay calificaciones
- TRANSPORTE DE ELECTRONES Y FOSFORILACION OXIDATIVAsegundoparcialpdfDocumento39 páginasTRANSPORTE DE ELECTRONES Y FOSFORILACION OXIDATIVAsegundoparcialpdfDebora RuizAún no hay calificaciones
- Receptores MuscarinicosDocumento12 páginasReceptores MuscarinicosMichell SchecterAún no hay calificaciones
- Dialnet CicatrizacionDeLesionSecundariaAAmputacionDeDedoIn 4609991Documento6 páginasDialnet CicatrizacionDeLesionSecundariaAAmputacionDeDedoIn 4609991Michell SchecterAún no hay calificaciones
- Práctica Determinación de ProteínasDocumento6 páginasPráctica Determinación de ProteínasMichell SchecterAún no hay calificaciones
- Práctica Titulación de SolucionesDocumento6 páginasPráctica Titulación de SolucionesMichell SchecterAún no hay calificaciones
- Kolin PlusDocumento2 páginasKolin PlusEdu LHAún no hay calificaciones
- Interpretacion y Comunicación CelularDocumento16 páginasInterpretacion y Comunicación CelularJhohan Andersson Sanchez Ylquimiche100% (1)
- Metabolismo de Compuestos Nitrogenados 01Documento56 páginasMetabolismo de Compuestos Nitrogenados 01Enzon BritoAún no hay calificaciones
- Amino AcidoDocumento40 páginasAmino AcidoJaime Lagos BorquezAún no hay calificaciones
- Ficha Tecnica - Stembolix BajaDocumento4 páginasFicha Tecnica - Stembolix BajaFABIAN GASTELUM LEYVAAún no hay calificaciones
- Clasificacion de Receptores RECEPTORES DE FÁRMACOSDocumento3 páginasClasificacion de Receptores RECEPTORES DE FÁRMACOSMerly SanchezAún no hay calificaciones
- Lectura Obligatoria de Semana 12Documento55 páginasLectura Obligatoria de Semana 12Mariabel Garcia CanalesAún no hay calificaciones
- 04-Proteínas y EnzimasDocumento20 páginas04-Proteínas y EnzimasAlberto Nsue Nzang AnguéAún no hay calificaciones
- Informe 1Documento10 páginasInforme 1guillermo malaver rodriguezAún no hay calificaciones
- Tema VitaminasDocumento11 páginasTema VitaminasHector Gpe Rangel RodriguezAún no hay calificaciones
- Taller BiomoleculasDocumento14 páginasTaller Biomoleculascristian bAún no hay calificaciones
- Actividad 1 - Evaluación Inicial - Evaluar Presaberes Bioquimica MetabolicaDocumento84 páginasActividad 1 - Evaluación Inicial - Evaluar Presaberes Bioquimica Metabolicaleonardo valencia100% (4)
- 2015-1 Bioquimica Gral. Prof. P. Woll Plan 2003Documento7 páginas2015-1 Bioquimica Gral. Prof. P. Woll Plan 2003DKNAún no hay calificaciones
- Ruta de Las Pentosas FosfatoDocumento26 páginasRuta de Las Pentosas FosfatoClaudio LeivaAún no hay calificaciones
- Mapa Conceptual Del MetabolismoDocumento2 páginasMapa Conceptual Del MetabolismoDiego Andres Garcia100% (1)
- Etapas de Replicación Viral.Documento23 páginasEtapas de Replicación Viral.baltazar flores100% (1)
- 10.regulación Génica en EucariotasDocumento20 páginas10.regulación Génica en EucariotasAgostina de MaríaAún no hay calificaciones
- Fisiologia EndocrinaDocumento39 páginasFisiologia EndocrinaTomas Garciaguirre100% (1)
- Metabolismo Lipidos PDFDocumento67 páginasMetabolismo Lipidos PDFarmando moralesAún no hay calificaciones
- PCR (Reacción en Cadena de La Polimerasa)Documento15 páginasPCR (Reacción en Cadena de La Polimerasa)Andrea MendozaAún no hay calificaciones
- Taller Viii Sem QcaDocumento4 páginasTaller Viii Sem QcamariaAún no hay calificaciones
- Sesión 11 (PROTEOGLICANOS)Documento24 páginasSesión 11 (PROTEOGLICANOS)Josver PretellAún no hay calificaciones
- Clasificascion de Monosacaridos CarbohidratosDocumento4 páginasClasificascion de Monosacaridos CarbohidratosRosario Estrada JacomeAún no hay calificaciones
- Las Vitaminas en La Alimentacion de CrustaceosDocumento5 páginasLas Vitaminas en La Alimentacion de Crustaceoskeyla yellicza espiritu guevaraAún no hay calificaciones
- Descubrimiento Del ARNDocumento5 páginasDescubrimiento Del ARNVane CastrejonAún no hay calificaciones
- Ciclo de KrebsDocumento2 páginasCiclo de KrebsMarjorie BelalcazarAún no hay calificaciones
- Entamoeba Histolytica EnzimasDocumento3 páginasEntamoeba Histolytica EnzimasDiana Valeria García ContrerasAún no hay calificaciones
- Quim4050 Cap4 EstudianteDocumento42 páginasQuim4050 Cap4 EstudianteNb + XB = AVAún no hay calificaciones
- Silabo de Bioquimica y Nutricion 2014-IiDocumento16 páginasSilabo de Bioquimica y Nutricion 2014-IiPercy RamírezAún no hay calificaciones
- Lípids Saponificables SimplesDocumento14 páginasLípids Saponificables SimplesJdjdjddAún no hay calificaciones