También podría gustarte
- Integración funcional del celador en el área de urgenciasDe EverandIntegración funcional del celador en el área de urgenciasAún no hay calificaciones
- Seis Argumentos A Favor de La Clonación Humana Que No Se SostienenDocumento6 páginasSeis Argumentos A Favor de La Clonación Humana Que No Se SostienenDkg AcostaAún no hay calificaciones
- Patogenia ViricaDocumento41 páginasPatogenia ViricafatycasAún no hay calificaciones
- Deficiencia Selectiva IgaDocumento12 páginasDeficiencia Selectiva IgaValenciaErikAún no hay calificaciones
- Replicación ViralDocumento25 páginasReplicación ViralAndrea ArdilaAún no hay calificaciones
- Tercer Parcial PATOLOGÍADocumento37 páginasTercer Parcial PATOLOGÍAVale Echavarri100% (1)
- Rebeca Lancefield Clasificación de EstreptococosDocumento6 páginasRebeca Lancefield Clasificación de EstreptococosBertha Lilia SalinasAún no hay calificaciones
- Aportes A La MicrobiologiaDocumento3 páginasAportes A La MicrobiologiaCaleb BuscagliaAún no hay calificaciones
- Casos Clínicos VirologíaDocumento8 páginasCasos Clínicos VirologíaARELY MURIEL MORENOAún no hay calificaciones
- Marco Teorico y Antecedentes-EthelDocumento22 páginasMarco Teorico y Antecedentes-EtheljeancAún no hay calificaciones
- CAPÍTULO 26 - EstómagoDocumento118 páginasCAPÍTULO 26 - Estómagodrifter527Aún no hay calificaciones
- Marcadores inmunológicos en diabetesDocumento4 páginasMarcadores inmunológicos en diabetesMaNuQlAún no hay calificaciones
- Fraccionamiento Celular Con Higado de VacaDocumento10 páginasFraccionamiento Celular Con Higado de VacaBraulio GarciaAún no hay calificaciones
- Principios de Asepsia Quirurgica y Atencion PrehospitalariaDocumento15 páginasPrincipios de Asepsia Quirurgica y Atencion PrehospitalariaBerenice Calero100% (1)
- Patogenia ViralDocumento25 páginasPatogenia Viralzthii_orthegaAún no hay calificaciones
- Bacterias de Importancia MédicaDocumento7 páginasBacterias de Importancia MédicaMoisés Salazar0% (1)
- Inmunidad InnataDocumento6 páginasInmunidad InnataDamaris Jetzabell Intriago ZamoraAún no hay calificaciones
- Infecciones OportunistasDocumento64 páginasInfecciones Oportunistasjrvg2007@gmail.comAún no hay calificaciones
- Autoevaluacion 2 PDFDocumento4 páginasAutoevaluacion 2 PDFBS3091Aún no hay calificaciones
- Vibrio Campylobacter y HelicobacterDocumento11 páginasVibrio Campylobacter y HelicobacterEliza ChanAún no hay calificaciones
- Ix Exploración Del Nervio GolosofaringeoDocumento2 páginasIx Exploración Del Nervio GolosofaringeoMohamed Bilal AbouAún no hay calificaciones
- Clase de SarconidosDocumento14 páginasClase de SarconidosTania PardoAún no hay calificaciones
- Cuestionario Rabia PolioDocumento4 páginasCuestionario Rabia PolioAaron Balbin CahuanaAún no hay calificaciones
- Etapa PrimariaDocumento5 páginasEtapa PrimariaEmérita Chaquila TineoAún no hay calificaciones
- Medicina Del AltiplanoDocumento32 páginasMedicina Del Altiplanodanitza pilco100% (1)
- Enterobiasis 131113161700 Phpapp01Documento17 páginasEnterobiasis 131113161700 Phpapp01Rafael valverde garro100% (1)
- CalcitoninaDocumento2 páginasCalcitoninaRodríguez Lovera Juan CarlosAún no hay calificaciones
- Caso Clínico HongosDocumento3 páginasCaso Clínico HongosBrandon DB100% (1)
- Parciales de HistologíaDocumento37 páginasParciales de HistologíaFranco CostaAún no hay calificaciones
- Inmunidad Frente A ParasitosDocumento17 páginasInmunidad Frente A ParasitosJose M. VillarroelAún no hay calificaciones
- Torax Anterior: 6ta Costal Reborde CostaDocumento1 páginaTorax Anterior: 6ta Costal Reborde CostaAna Cecilia MantillaAún no hay calificaciones
- Desmielinizacion PontinaDocumento6 páginasDesmielinizacion PontinagilbertochapaAún no hay calificaciones
- Vigilancia Inmunológica A Inmunidad AntitumoralDocumento11 páginasVigilancia Inmunológica A Inmunidad AntitumoralSher Valerio100% (1)
- Respuestas Parcial de Microbiología 3, VirusDocumento20 páginasRespuestas Parcial de Microbiología 3, VirusDavid Ruiz RuizAún no hay calificaciones
- Semana 1 PDFDocumento27 páginasSemana 1 PDFJose galvezAún no hay calificaciones
- Parasitología: Enfermedades causadas por agentes biológicosDocumento85 páginasParasitología: Enfermedades causadas por agentes biológicosAlfredo Benjamin Fuentes OrtizAún no hay calificaciones
- Observación microorganismos alimentosDocumento2 páginasObservación microorganismos alimentosGabriela Pasion BazanAún no hay calificaciones
- Holcomb y Ashcraft Cirugía Pediátrica - 7 Edición-676-686Documento11 páginasHolcomb y Ashcraft Cirugía Pediátrica - 7 Edición-676-686Brandon ChavarriaAún no hay calificaciones
- Historia de La Prevención de EnfermedadesDocumento26 páginasHistoria de La Prevención de EnfermedadesDani CicardiniAún no hay calificaciones
- Analisis de Leviticos Relacionados Con Salud PublicaDocumento7 páginasAnalisis de Leviticos Relacionados Con Salud PublicaSteven Plaza100% (1)
- ENTOMOFTOROMICOSISDocumento6 páginasENTOMOFTOROMICOSISMaritza Ortiz100% (1)
- Capítulo 4 Obstetricia de WilliamsDocumento24 páginasCapítulo 4 Obstetricia de WilliamsAlejandra Vera AlvarezAún no hay calificaciones
- Sifilis CongenitaDocumento24 páginasSifilis CongenitaCarlos ChavezAún no hay calificaciones
- Flora Microbiana Normal - MurrayDocumento5 páginasFlora Microbiana Normal - MurrayMARIAMNE AZUCENA IBARRA RESENDIZAún no hay calificaciones
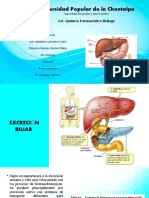
- Excreción BiliarDocumento10 páginasExcreción BiliarFranx SanchezAún no hay calificaciones
- Salmonella Shigella V Cholerae y E ColiDocumento14 páginasSalmonella Shigella V Cholerae y E ColiEmiliano MasseroniAún no hay calificaciones
- Laboratorio 1 Biologia MolecularDocumento6 páginasLaboratorio 1 Biologia MolecularJenny P. de YelmoAún no hay calificaciones
- 8 - HipersensibilidadDocumento42 páginas8 - HipersensibilidadCarlos Edwin Perez Garcia DraptekAún no hay calificaciones
- Determinacion de Proteinas PDFDocumento77 páginasDeterminacion de Proteinas PDFKarla Polo PiminchumoAún no hay calificaciones
- Angiostrongiliasis ParagonimiasisDocumento22 páginasAngiostrongiliasis ParagonimiasisMaria Jose Jimenez100% (1)
- El Hígado (Semiologia)Documento24 páginasEl Hígado (Semiologia)Enrique Ollarbe Castillo100% (1)
- Sindrome de Swyer - Wikipedia, La Enciclopedia LibreDocumento3 páginasSindrome de Swyer - Wikipedia, La Enciclopedia LibreAlejandro VargasAún no hay calificaciones
- Cultivo hongos dermatofitosDocumento7 páginasCultivo hongos dermatofitosKei QPAún no hay calificaciones
- MENINGITISDocumento16 páginasMENINGITISSandra DicttoAún no hay calificaciones
- Cancer Uterino Peru CervixDocumento10 páginasCancer Uterino Peru CervixGigi Ramos Galarza100% (1)
- ARENAVIRUS Junin MachupoDocumento11 páginasARENAVIRUS Junin Machupoelnegro300Aún no hay calificaciones
- Expo SIDA: Origen, propagación y síntomas del VIH/SIDADocumento2 páginasExpo SIDA: Origen, propagación y síntomas del VIH/SIDAXochitl PerezAún no hay calificaciones
- Proyecto Parasito Mugil Cpehalus FinalDocumento7 páginasProyecto Parasito Mugil Cpehalus FinalAlexis DíazAún no hay calificaciones
- Adriana Fiorella Medranda Pin-virologia-7-A Consulta 2Documento12 páginasAdriana Fiorella Medranda Pin-virologia-7-A Consulta 2greco menesesAún no hay calificaciones
- Unidad 2 CompendioDocumento14 páginasUnidad 2 CompendioAlejandro BenvenutaAún no hay calificaciones
- Ubicación e instalación de apiarios para producción apícolaDocumento6 páginasUbicación e instalación de apiarios para producción apícolabaltazar floresAún no hay calificaciones
- Estructuras FúngicasDocumento52 páginasEstructuras Fúngicasbaltazar floresAún no hay calificaciones
- Elaboración de Concentrados.Documento2 páginasElaboración de Concentrados.baltazar floresAún no hay calificaciones
- Producción y Comercio de Ganado y Carne de Bovino en El MundoDocumento3 páginasProducción y Comercio de Ganado y Carne de Bovino en El Mundobaltazar floresAún no hay calificaciones
- Criterios de selección de la carne bovinaDocumento4 páginasCriterios de selección de la carne bovinabaltazar floresAún no hay calificaciones
- Análisis de VideoDocumento6 páginasAnálisis de Videobaltazar floresAún no hay calificaciones
- Álbum de Razas de EquinosDocumento24 páginasÁlbum de Razas de Equinosbaltazar flores100% (2)
- Libro de InsectosDocumento277 páginasLibro de InsectosEstefani BejarAún no hay calificaciones
- Universidad Realística de México: Sarna sarcóptica en caninosDocumento3 páginasUniversidad Realística de México: Sarna sarcóptica en caninosbaltazar floresAún no hay calificaciones
- Gatos y EnfermedadesDocumento16 páginasGatos y Enfermedadesbaltazar floresAún no hay calificaciones
- Cera de Abejas.Documento6 páginasCera de Abejas.baltazar floresAún no hay calificaciones
- Examen NeurológicoDocumento9 páginasExamen Neurológicobaltazar floresAún no hay calificaciones
- Cuadros Sinópticos de Enfermedades.Documento9 páginasCuadros Sinópticos de Enfermedades.baltazar floresAún no hay calificaciones
- Familias de Virus.Documento8 páginasFamilias de Virus.baltazar floresAún no hay calificaciones
- Fisiología Animal IIDocumento141 páginasFisiología Animal IITatiana VelascoAún no hay calificaciones
- 2019 Freemartinismo Ganado BovinoDocumento18 páginas2019 Freemartinismo Ganado BovinoKarla Areli VFAún no hay calificaciones
- Importancia de los minerales en la nutrición animalDocumento4 páginasImportancia de los minerales en la nutrición animalbaltazar floresAún no hay calificaciones
- Cultivo de Microorganismos UrmDocumento46 páginasCultivo de Microorganismos Urmbaltazar floresAún no hay calificaciones
- Protocolo McMasterDocumento4 páginasProtocolo McMasterbaltazar floresAún no hay calificaciones
- Ciclo MenstrualDocumento4 páginasCiclo Menstrualbaltazar floresAún no hay calificaciones
- Diabetes canina resumenDocumento3 páginasDiabetes canina resumenbaltazar floresAún no hay calificaciones
- Paradigmas de La Ciencia.Documento4 páginasParadigmas de La Ciencia.baltazar floresAún no hay calificaciones
- Baño GeneralDocumento11 páginasBaño Generalbaltazar floresAún no hay calificaciones
- Aplomos Actividad 1Documento2 páginasAplomos Actividad 1baltazar floresAún no hay calificaciones
- Antidiarreicos Veterinarios URMDocumento12 páginasAntidiarreicos Veterinarios URMbaltazar floresAún no hay calificaciones
- Aplomos Actividad 1Documento2 páginasAplomos Actividad 1baltazar floresAún no hay calificaciones
- Anatomía Externa e Interna de La Abeja - TóraxDocumento9 páginasAnatomía Externa e Interna de La Abeja - Tóraxbaltazar floresAún no hay calificaciones
- Anatomía MDocumento9 páginasAnatomía Mbaltazar floresAún no hay calificaciones
- Alimentos Tóxicos para Pequeñas EspeciesDocumento8 páginasAlimentos Tóxicos para Pequeñas Especiesbaltazar floresAún no hay calificaciones
- HerpesDocumento33 páginasHerpesSaraith PozosAún no hay calificaciones
- Paso La VillaDocumento38 páginasPaso La Villaluis toscanoAún no hay calificaciones
- Esquema de Vacunación, 2018Documento1 páginaEsquema de Vacunación, 2018Stefany Guardado100% (1)
- Microbiología Ii CorregidoDocumento315 páginasMicrobiología Ii CorregidoHenry Palomino GutierrezAún no hay calificaciones
- Algoritmo Exantemas Vesiculo-AmpollososDocumento3 páginasAlgoritmo Exantemas Vesiculo-AmpollososAngel GUAún no hay calificaciones
- Dmitri Ivanovski biografíaDocumento3 páginasDmitri Ivanovski biografíaWilder Jean Patricio MartinezAún no hay calificaciones
- 21 Taxonomia ViralDocumento24 páginas21 Taxonomia ViralAurico Sousa Fonseca85% (13)
- Fiebre Amarilla (YF)Documento12 páginasFiebre Amarilla (YF)Kyara TamaraAún no hay calificaciones
- TAMIFLUDocumento2 páginasTAMIFLUSheila RabanalAún no hay calificaciones
- Varicela HemorragicaDocumento12 páginasVaricela HemorragicaandyAún no hay calificaciones
- VIRUSDocumento15 páginasVIRUSClara01 wenAún no hay calificaciones
- Práctica N 13. Diagnóstico COVID-19 - SBDocumento25 páginasPráctica N 13. Diagnóstico COVID-19 - SBNicole Mireya Aleman ChaconAún no hay calificaciones
- PPT. Covid 19 Salesiano PDFDocumento20 páginasPPT. Covid 19 Salesiano PDFAlex HQAún no hay calificaciones
- DengueDocumento5 páginasDenguepretylislalkkAún no hay calificaciones
- Virus Arn Bunya-Arena (Autoguardado)Documento43 páginasVirus Arn Bunya-Arena (Autoguardado)ClauMcClauAún no hay calificaciones
- 23.formato Informe Movimiento Biologico 2021Documento40 páginas23.formato Informe Movimiento Biologico 2021Liris Del Carmen Berrocal PlazasAún no hay calificaciones
- Introducción y cierre de textos sobre aislamiento social por covid-19Documento2 páginasIntroducción y cierre de textos sobre aislamiento social por covid-19Carlos Alvarado100% (3)
- Diferencias Entre Varicela y SD Mano, Pie, BocaDocumento1 páginaDiferencias Entre Varicela y SD Mano, Pie, BocaCamilita Ponce 1109Aún no hay calificaciones
- cuestionario-del-modulo-2-revision-del-intento-2Documento22 páginascuestionario-del-modulo-2-revision-del-intento-2Nathaly ChoezAún no hay calificaciones
- Misael Emilio Martinez Gonzalez: Sars-Cov-2 (2019-Ncov) PCRDocumento1 páginaMisael Emilio Martinez Gonzalez: Sars-Cov-2 (2019-Ncov) PCRMenisael MartAún no hay calificaciones
- Enfermedades Respiratorias ViralesDocumento72 páginasEnfermedades Respiratorias ViralesSebastian Quintero MontealegreAún no hay calificaciones
- Plan de La Estrategia Sanitaria de ZoonosisDocumento1 páginaPlan de La Estrategia Sanitaria de ZoonosisMarco MarcoAún no hay calificaciones
- Lineas CelularesDocumento5 páginasLineas CelularesAnthony Merino Estrella50% (2)
- Trabajo Terminado Sarampion.Documento13 páginasTrabajo Terminado Sarampion.Gerson LopezAún no hay calificaciones
- E123 Clavijo+Naula+et+alDocumento18 páginasE123 Clavijo+Naula+et+alSantiago GomezAún no hay calificaciones
- Influenza Equina-ToMÁS MERÁZDocumento2 páginasInfluenza Equina-ToMÁS MERÁZRamsés MadiAún no hay calificaciones
- Infografía COVID 19 y VacunasDocumento1 páginaInfografía COVID 19 y VacunasDiana GiselleAún no hay calificaciones
- Antivirales PresentacionDocumento43 páginasAntivirales PresentacionLourdes Ortiz OchoaAún no hay calificaciones
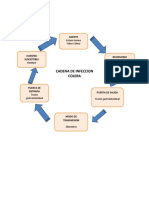
- Cadena de Infeccion EnfermedadesDocumento9 páginasCadena de Infeccion EnfermedadesDinel MolinaAún no hay calificaciones
- Choice VirusDocumento105 páginasChoice VirusemilyAún no hay calificaciones
- Genética general: Libro de textoDe EverandGenética general: Libro de textoCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- TDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosDe EverandTDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosCalificación: 4 de 5 estrellas4/5 (8)
- Psiconeuroinmunología para la práctica clínicaDe EverandPsiconeuroinmunología para la práctica clínicaCalificación: 5 de 5 estrellas5/5 (4)
- En busca de la mente: El largo camino para comprender la vida mental (y lo que aún queda por descubrir)De EverandEn busca de la mente: El largo camino para comprender la vida mental (y lo que aún queda por descubrir)Calificación: 4.5 de 5 estrellas4.5/5 (3)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Ansiedad infantil. Los trastornos explicados a los padresDe EverandAnsiedad infantil. Los trastornos explicados a los padresCalificación: 4.5 de 5 estrellas4.5/5 (25)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Abrázame fuerte: Siete conversaciones para un amor duraderoDe EverandAbrázame fuerte: Siete conversaciones para un amor duraderoManu BerásteguiCalificación: 4.5 de 5 estrellas4.5/5 (13)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- Trauma, miedo y amor: Hacia una autonomía interior con la ayuda de las constelacionesDe EverandTrauma, miedo y amor: Hacia una autonomía interior con la ayuda de las constelacionesCalificación: 1 de 5 estrellas1/5 (1)
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- El libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)De EverandEl libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)Calificación: 3 de 5 estrellas3/5 (2)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- Póngase En Forma Sin Salir De Su CasaDe EverandPóngase En Forma Sin Salir De Su CasaCalificación: 4.5 de 5 estrellas4.5/5 (4)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- El autismo: Reflexiones y pautas para comprenderlo y abordarloDe EverandEl autismo: Reflexiones y pautas para comprenderlo y abordarloCalificación: 4 de 5 estrellas4/5 (7)
- Terapia de vidas pasadas: Un camino hacia la luz del alma. Técnica y prácticaDe EverandTerapia de vidas pasadas: Un camino hacia la luz del alma. Técnica y prácticaCalificación: 4.5 de 5 estrellas4.5/5 (11)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Prescripción de ejercico físico para la saludDe EverandPrescripción de ejercico físico para la saludCalificación: 5 de 5 estrellas5/5 (1)