También podría gustarte
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Sistema nervioso: Neuroanatomía funcional y clínicaDe EverandSistema nervioso: Neuroanatomía funcional y clínicaCalificación: 4 de 5 estrellas4/5 (4)
- Enrocados de ProtecciónDocumento25 páginasEnrocados de ProtecciónPedro Bartolome100% (3)
- Que Disciplinas Forman Parte de La Neurociencia Del LenguajeDocumento3 páginasQue Disciplinas Forman Parte de La Neurociencia Del LenguajeNANO perezAún no hay calificaciones
- Manual de prácticas de Neuroanatomía: Laboratorio de morfologíaDe EverandManual de prácticas de Neuroanatomía: Laboratorio de morfologíaAún no hay calificaciones
- Dominancia HemisfericaDocumento8 páginasDominancia HemisfericaCarlos Alfredo Castillo Delgado100% (1)
- Dominancia Hemisferica Art Med.Documento8 páginasDominancia Hemisferica Art Med.Sabrina YanceAún no hay calificaciones
- Articulo RadiologiaDocumento9 páginasArticulo RadiologiaNata VMAún no hay calificaciones
- Haro - 2022 - Potenciales Evocados y LenguajeDocumento20 páginasHaro - 2022 - Potenciales Evocados y LenguajeMicaela DiAún no hay calificaciones
- Resumen Neurociencia Del LenguajeDocumento6 páginasResumen Neurociencia Del LenguajeNuria FernandezAún no hay calificaciones
- Neurolingüística y Programacion LingüísticaDocumento23 páginasNeurolingüística y Programacion LingüísticaRukia Nair Fernandez PaniaguaAún no hay calificaciones
- Modelos Neuroanatomo FuncionalesDocumento21 páginasModelos Neuroanatomo FuncionalesAyelen ChaconAún no hay calificaciones
- Leal Catalán Patricia UD3 Estudio de CasoDocumento3 páginasLeal Catalán Patricia UD3 Estudio de CasoPatri LCAún no hay calificaciones
- Reorganizacion LgjeDocumento4 páginasReorganizacion LgjeCony GCAún no hay calificaciones
- Mapa Cortical y SubcorticalDocumento9 páginasMapa Cortical y SubcorticalAndreaAún no hay calificaciones
- NeurolingüísticaDocumento7 páginasNeurolingüísticaDaniel Henriquez100% (1)
- Area Cerebral Del Lenguaje, Una Reconsideracion FuncionalDocumento10 páginasArea Cerebral Del Lenguaje, Una Reconsideracion FuncionalBegoña MartinezAún no hay calificaciones
- Marrero Aguiar - 2022 - Neuroimágenes y LenguajeDocumento25 páginasMarrero Aguiar - 2022 - Neuroimágenes y LenguajeMicaela DiAún no hay calificaciones
- Qué Hay de Nuevo en Los Métodos de NeuroimagenDocumento24 páginasQué Hay de Nuevo en Los Métodos de Neuroimagenicaina.jgAún no hay calificaciones
- Programas para Análisis de Señales y Evaluación Cognoscitiva - GuevaraDocumento180 páginasProgramas para Análisis de Señales y Evaluación Cognoscitiva - GuevaraLuis PrietoAún no hay calificaciones
- RESUMENDocumento16 páginasRESUMENBianca Teresa Meillon GarciaAún no hay calificaciones
- M28T4 Alteraciones Motoras y Del LenguajeDocumento12 páginasM28T4 Alteraciones Motoras y Del LenguajesusanacastrodsAún no hay calificaciones
- MRI Lenguaje y Audicion (2014)Documento35 páginasMRI Lenguaje y Audicion (2014)Olivia Grasty VillanuevaAún no hay calificaciones
- Circuitos Motores Del HablaDocumento17 páginasCircuitos Motores Del HablaLidaConstanzaMendezAún no hay calificaciones
- Coorrelación Anatomofuncional de LaafasiaDocumento18 páginasCoorrelación Anatomofuncional de LaafasiaAldo Hip NaranjoAún no hay calificaciones
- Resonancia Magnética Funcional. AscencioDocumento5 páginasResonancia Magnética Funcional. AscencioPAULA ANDREA NUÑEZ ORTIZAún no hay calificaciones
- Logopedia en Paciente Con Mapeo Corticalintraoperatorio.Documento7 páginasLogopedia en Paciente Con Mapeo Corticalintraoperatorio.Estefania OrtuñoAún no hay calificaciones
- Capitulo IV Afasias Tipologia Clinico TopograficaDocumento23 páginasCapitulo IV Afasias Tipologia Clinico TopograficaJuan José Quesada Revelo100% (2)
- Ficha de Revisión de Artículo Empírico-1Documento3 páginasFicha de Revisión de Artículo Empírico-1ERIN HADIR CALDONAún no hay calificaciones
- Aplicaciones de La Resonancia Magnética Funcional en Pacientes Prequirúrgicos Funciones Motora, de Memoria y LingüísticaDocumento12 páginasAplicaciones de La Resonancia Magnética Funcional en Pacientes Prequirúrgicos Funciones Motora, de Memoria y LingüísticaClaudio López SepúlvedaAún no hay calificaciones
- Español Electromiografia para Hbala y DegluciónDocumento16 páginasEspañol Electromiografia para Hbala y Degluciónvaleria posadaAún no hay calificaciones
- Introducción A La RMF (Lisandro)Documento4 páginasIntroducción A La RMF (Lisandro)Ñaco Empanadas Libertador San MartinAún no hay calificaciones
- AfasiologiaDocumento1 páginaAfasiologiaJuanRAún no hay calificaciones
- Bases Neuroanatómicas Del LenguajeDocumento108 páginasBases Neuroanatómicas Del LenguajeBorja Fernández LópezAún no hay calificaciones
- El Enfoque de La Neurociencia CognitivaDocumento3 páginasEl Enfoque de La Neurociencia CognitivaAstrid Gallegos100% (1)
- Dislexia TresDocumento6 páginasDislexia TresSusanne MariAún no hay calificaciones
- El Sistema Lexical - Caramazza 1988 PDFDocumento16 páginasEl Sistema Lexical - Caramazza 1988 PDFginger_2011Aún no hay calificaciones
- TP Nº2 de Procesos Biologicos (2FINAL)Documento9 páginasTP Nº2 de Procesos Biologicos (2FINAL)Pía MartinAún no hay calificaciones
- Neuropsicologia - AfasiasDocumento9 páginasNeuropsicologia - AfasiasPichon RomAún no hay calificaciones
- Neuroimagen Funcional y LenguajeDocumento9 páginasNeuroimagen Funcional y LenguajeNan JimenezAún no hay calificaciones
- Neuroanatomía Del LenguajeDocumento4 páginasNeuroanatomía Del LenguajeHector Agullo RiveroAún no hay calificaciones
- Secid 2012 PDFDocumento229 páginasSecid 2012 PDFEmperor52Aún no hay calificaciones
- Estudios de Neurodiagnostico Del HablaDocumento5 páginasEstudios de Neurodiagnostico Del HablaPaola BrimAún no hay calificaciones
- Red Neruonal de Las AfasiasDocumento12 páginasRed Neruonal de Las AfasiasMilenaAún no hay calificaciones
- Neuroimagen Funcional y LenguajeDocumento10 páginasNeuroimagen Funcional y LenguajeFERNANDO YUMBO TACURIAún no hay calificaciones
- Neurociencias Cognitivas - Cap.19 Lenguaje 1Documento29 páginasNeurociencias Cognitivas - Cap.19 Lenguaje 1isa vAún no hay calificaciones
- Aspectos-Neuropsicologicos RMNFDocumento5 páginasAspectos-Neuropsicologicos RMNFJulia LemboAún no hay calificaciones
- Utilidad Clínica Del Electroencefalograma en El Trastorno Por Déficit de Atención Con Hiperactividad - Una Actualización de La InvestigaciónDocumento34 páginasUtilidad Clínica Del Electroencefalograma en El Trastorno Por Déficit de Atención Con Hiperactividad - Una Actualización de La InvestigaciónDavid De Luna MartínezAún no hay calificaciones
- El Modelo DIVADocumento21 páginasEl Modelo DIVAElianaAún no hay calificaciones
- Notas ClaseDocumento15 páginasNotas ClaseSara Rodriguez MolinaAún no hay calificaciones
- Localizacion Cerebral de Las Funciones EjecutivasDocumento5 páginasLocalizacion Cerebral de Las Funciones EjecutivasJavi VeraAún no hay calificaciones
- Fun C I Ones Audit I Vas Cerebral EsDocumento12 páginasFun C I Ones Audit I Vas Cerebral Eskiratau2000Aún no hay calificaciones
- Resonancia MagnéticaDocumento16 páginasResonancia MagnéticaMar CFAún no hay calificaciones
- Acceso Ultra Rápido A Las Palabras en La Afasia CrónicaDocumento13 páginasAcceso Ultra Rápido A Las Palabras en La Afasia CrónicaNixza Vera VergaraAún no hay calificaciones
- Tarea 4 de PsicolinguisticaDocumento5 páginasTarea 4 de PsicolinguisticaLuisa PerdomoAún no hay calificaciones
- RAE # - (2 Páginas) Estudiantes: ID:: María Paula Aguas Ochoa y Melany Valero 000482161 - 000463291Documento2 páginasRAE # - (2 Páginas) Estudiantes: ID:: María Paula Aguas Ochoa y Melany Valero 000482161 - 000463291Melany Valero ZuletaAún no hay calificaciones
- Las Neurociencias Cognitivas Han Tenido Un Desarrollo Formidable en Los Últimos AñosDocumento1 páginaLas Neurociencias Cognitivas Han Tenido Un Desarrollo Formidable en Los Últimos AñosCristel CarpioAún no hay calificaciones
- Morfología del sistema nervioso: Parte 1: médula espinal y encéfaloDe EverandMorfología del sistema nervioso: Parte 1: médula espinal y encéfaloAún no hay calificaciones
- Manual de prácticas de Neuroanatomía 2da edición: Laboratorio de morfologíaDe EverandManual de prácticas de Neuroanatomía 2da edición: Laboratorio de morfologíaAún no hay calificaciones
- La fonética forense: Nuevos retos y nuevas líneas de investigaciónDe EverandLa fonética forense: Nuevos retos y nuevas líneas de investigaciónAún no hay calificaciones
- CIE10Documento5 páginasCIE10ceballosbisAún no hay calificaciones
- CIE10Documento5 páginasCIE10ceballosbisAún no hay calificaciones
- Recursos Tecnologicos para La Intervencion Del Lenguaje Oral y EscritoDocumento13 páginasRecursos Tecnologicos para La Intervencion Del Lenguaje Oral y EscritoceballosbisAún no hay calificaciones
- EL NIÑO TEL EN LA ESCUELA Gerardo AguadoDocumento8 páginasEL NIÑO TEL EN LA ESCUELA Gerardo Aguadoceballosbis100% (2)
- Habilitación FuncionalDocumento6 páginasHabilitación FuncionalceballosbisAún no hay calificaciones
- Estrategias de Intervencion en Los TEL de Educacion InfantilDocumento17 páginasEstrategias de Intervencion en Los TEL de Educacion Infantilceballosbis100% (1)
- Primera Noticia At. TempranaDocumento46 páginasPrimera Noticia At. TempranaGarbayoAún no hay calificaciones
- Escolarizacion Niños TELDocumento4 páginasEscolarizacion Niños TELceballosbisAún no hay calificaciones
- Libro Trastornos Del Lenguaje en La InfanciaDocumento10 páginasLibro Trastornos Del Lenguaje en La Infanciaceballosbis100% (2)
- Trastornos de La Comunicación y El LenguajeDocumento41 páginasTrastornos de La Comunicación y El Lenguajedaniel_ledesma_24100% (5)
- Intervencion Familiar en Niños Con TLDocumento29 páginasIntervencion Familiar en Niños Con TLceballosbisAún no hay calificaciones
- Trastornos Del Lenguaje Dra Rosa Gassio-SubirachsDocumento5 páginasTrastornos Del Lenguaje Dra Rosa Gassio-Subirachsceballosbis100% (2)
- AFATEL Ficha de InscripcionDocumento2 páginasAFATEL Ficha de InscripcionceballosbisAún no hay calificaciones
- Manual para Padres de Niños TELDocumento20 páginasManual para Padres de Niños TELceballosbis0% (1)
- Desarrollo Temprano, Autismo y Trastornos Del Lenguaje (J.Martos)Documento8 páginasDesarrollo Temprano, Autismo y Trastornos Del Lenguaje (J.Martos)info-TEAAún no hay calificaciones
- TELDocumento7 páginasTELJavi MoranteAún no hay calificaciones
- Entrevista Gerardo AguadoDocumento9 páginasEntrevista Gerardo Aguadoceballosbis100% (1)
- El Lenguaje Del Nio y Sus TrastornosDocumento12 páginasEl Lenguaje Del Nio y Sus TrastornosTammy DonosoAún no hay calificaciones
- El Pais Gerardo Aguado Especialista en Psicologia Del LenguajeDocumento3 páginasEl Pais Gerardo Aguado Especialista en Psicologia Del LenguajeceballosbisAún no hay calificaciones
- Indicadores Tempranos de Los TDLDocumento12 páginasIndicadores Tempranos de Los TDLceballosbis100% (2)
- M Monfort Escolarizacion TelDocumento3 páginasM Monfort Escolarizacion TelceballosbisAún no hay calificaciones
- Esperanza y Utopia Ernst Bloch Desde Ame PDFDocumento48 páginasEsperanza y Utopia Ernst Bloch Desde Ame PDFLarry DelaoAún no hay calificaciones
- Cema PR Fa12Documento10 páginasCema PR Fa12ReneVanegasPoloAún no hay calificaciones
- Repertorios Bibliográficos TrabajoDocumento8 páginasRepertorios Bibliográficos TrabajoJOSE PEROZAAún no hay calificaciones
- Propuestas de Escritura CreativaDocumento10 páginasPropuestas de Escritura CreativabiblioguadalentinAún no hay calificaciones
- RESUMEN DEL LIBRO - Quien Se Ha Llevado Mi QuesoDocumento3 páginasRESUMEN DEL LIBRO - Quien Se Ha Llevado Mi QuesoViviana SayasAún no hay calificaciones
- CosoDocumento50 páginasCosoFreddy Mauricio RamosAún no hay calificaciones
- Manual CTO CardiologiaDocumento2 páginasManual CTO CardiologiaElaine CarvalhoAún no hay calificaciones
- Factores de Cambio SocialDocumento3 páginasFactores de Cambio SocialDany LHAún no hay calificaciones
- Inteligencias Multiples y Aprendizaje IntegralDocumento9 páginasInteligencias Multiples y Aprendizaje Integraladriana machadoAún no hay calificaciones
- Poesia GuatemaltecaDocumento14 páginasPoesia GuatemaltecaRosmeryAún no hay calificaciones
- Numeros ComplejosDocumento4 páginasNumeros ComplejosLuis LzoAún no hay calificaciones
- Seis Ideas Falsas Sobre La GlobalizacionDocumento3 páginasSeis Ideas Falsas Sobre La GlobalizacionLuis Fernando SanclementeAún no hay calificaciones
- La Agonia de Rasu Niti de Jose Maria ArguedasDocumento4 páginasLa Agonia de Rasu Niti de Jose Maria Arguedasgaby0% (1)
- Evaluacion Final - Escenario 8ESTADISTICA INFERENCIALDocumento12 páginasEvaluacion Final - Escenario 8ESTADISTICA INFERENCIALStefania BolivarAún no hay calificaciones
- Intemperismo PDFDocumento46 páginasIntemperismo PDFvampiresangrienta0% (1)
- Estructura Social y Economica de Los MayasDocumento8 páginasEstructura Social y Economica de Los MayasIsrael MpAún no hay calificaciones
- 1er Parcial Unidad 3 Auditoria FinancieraDocumento20 páginas1er Parcial Unidad 3 Auditoria FinancieraBayron BeckerAún no hay calificaciones
- Orar Desde La PalabraDocumento20 páginasOrar Desde La PalabraDiácono Anderson Santana CunhaAún no hay calificaciones
- Guia 1 Algebra y FuncionesDocumento2 páginasGuia 1 Algebra y FuncionesOscar AcevedoAún no hay calificaciones
- Las Teorías Psicoanalíticas de La CreatividadDocumento5 páginasLas Teorías Psicoanalíticas de La CreatividadgigiAún no hay calificaciones
- Respuesta Metabólica Al TraumaDocumento22 páginasRespuesta Metabólica Al TraumaTania RodriguezAún no hay calificaciones
- Deposito de Inercia Enfriadoras HitopDocumento3 páginasDeposito de Inercia Enfriadoras HitopJuan Carlos SerranoAún no hay calificaciones
- SESION Porcentaje Areas m2Documento8 páginasSESION Porcentaje Areas m2Zolo C K Nada CAún no hay calificaciones
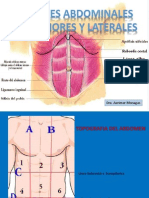
- Paredes AbdominalesDocumento57 páginasParedes Abdominalesaurimarmonagas100% (2)
- Técnicas Narrativas Tarea PsicometriaDocumento3 páginasTécnicas Narrativas Tarea PsicometriaJohn Carlos Rojas UrdánigoAún no hay calificaciones
- ConclusiónDocumento2 páginasConclusiónmonsecatalina85% (13)
- Guia Lengua y LiteraturaDocumento742 páginasGuia Lengua y LiteraturaMaria Magdalena Castro SánchezAún no hay calificaciones
- ANAMNESISDocumento2 páginasANAMNESISfire pyAún no hay calificaciones
- Aritmética - Semana 3 - 5.º AñoDocumento2 páginasAritmética - Semana 3 - 5.º Añojosé denilson herrera salasAún no hay calificaciones