También podría gustarte
- CarbohidratosDocumento28 páginasCarbohidratosAnonimo22100% (2)
- Ciclo de La PentosaDocumento46 páginasCiclo de La PentosaLuis Contreras ArtuzAún no hay calificaciones
- GlucólisisDocumento52 páginasGlucólisisEduardo Limay Coronado100% (5)
- Bioquimica - LipidosDocumento91 páginasBioquimica - LipidosNadia Torres100% (2)
- LipidosDocumento33 páginasLipidosJunior HernándezAún no hay calificaciones
- LIPIDOSDocumento69 páginasLIPIDOSVeronicaGonzalezAún no hay calificaciones
- Metabolismo de Lipidos ClaseDocumento16 páginasMetabolismo de Lipidos ClaseAndrea ManziniAún no hay calificaciones
- GlucolisisDocumento56 páginasGlucolisisXavi EscobarAún no hay calificaciones
- Metabolismo de CarbohidratosDocumento23 páginasMetabolismo de CarbohidratosJoss Spigut100% (2)
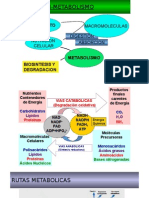
- Rutas MetabolicasDocumento35 páginasRutas Metabolicassam100% (1)
- Duplicación Del AdnDocumento12 páginasDuplicación Del AdnMariAún no hay calificaciones
- Vitaminas y MineralesDocumento34 páginasVitaminas y MineralesJ Carlos Meza CalderonAún no hay calificaciones
- Metabolismo de AminoacidosDocumento11 páginasMetabolismo de Aminoacidosapi-3696656100% (3)
- AMINOÁCIDOSDocumento9 páginasAMINOÁCIDOSMary Marquez GutierrezAún no hay calificaciones
- ProteínasDocumento22 páginasProteínasaideetoledo18100% (2)
- Metabolismo de LipidosDocumento99 páginasMetabolismo de LipidosANGUIE ANETTE MAMANI SUCASAIREAún no hay calificaciones
- Recursos para la enseñanza/aprendizaje del metabolismoDe EverandRecursos para la enseñanza/aprendizaje del metabolismoAún no hay calificaciones
- GLUCONEOGENESISDocumento32 páginasGLUCONEOGENESISzatanna66100% (2)
- Grasas y LípidosDocumento26 páginasGrasas y LípidosAlonsoAún no hay calificaciones
- GlucogenesisDocumento7 páginasGlucogenesisJareth Caldera100% (3)
- Ciclo de KrebsDocumento25 páginasCiclo de KrebsJasonRodriguezGuerra100% (2)
- Metabolismo de Los Carbohidratos ExpoDocumento35 páginasMetabolismo de Los Carbohidratos ExpoJoelTevesMendoza100% (2)
- Metabolismo de Los CarbohidratosDocumento51 páginasMetabolismo de Los CarbohidratosMaria Angelica Santillan Carrasco0% (1)
- Metabolismo de CarbohidratosDocumento52 páginasMetabolismo de CarbohidratosLauraSofiaRamirezAún no hay calificaciones
- Pentosas FosfatoDocumento21 páginasPentosas FosfatoSlovan Ochoa100% (1)
- 8 Metabolismo de LipidosDocumento52 páginas8 Metabolismo de Lipidosalexandra100% (2)
- Metabolismo de Los CarbohidratosDocumento117 páginasMetabolismo de Los CarbohidratosAscrusAún no hay calificaciones
- Metabolismo de LípidosDocumento68 páginasMetabolismo de LípidossayhandalAún no hay calificaciones
- B-Oxidacion y Sintesis de Acidos GrasosDocumento22 páginasB-Oxidacion y Sintesis de Acidos GrasosPatricio Coronado75% (8)
- GlucogenolisisDocumento15 páginasGlucogenolisisTammy Carrillo Recabarren100% (1)
- GluconeogénesisDocumento18 páginasGluconeogénesisSergio Castillo100% (3)
- Rutas MetabólicasDocumento48 páginasRutas MetabólicasEduardo Aguilar beltranAún no hay calificaciones
- GlucolisisDocumento26 páginasGlucolisisAle JoyaAún no hay calificaciones
- Fosforilación OxidativaDocumento25 páginasFosforilación OxidativaEmilio Jeanpaul Mitacc GutierrezAún no hay calificaciones
- 7 LipidosDocumento89 páginas7 Lipidosrafael100% (1)
- Integracion MetabolicaDocumento53 páginasIntegracion MetabolicaIbelise Gonzalez67% (3)
- GlucolisisDocumento15 páginasGlucolisisalejo_1994244644100% (1)
- Examen de BioquimicaDocumento2 páginasExamen de BioquimicaYuMi BautistaAún no hay calificaciones
- ¿Ha Conseguido El Dr. Marty Hinz Curar Los Sintomas Del ParkinsonDocumento9 páginas¿Ha Conseguido El Dr. Marty Hinz Curar Los Sintomas Del ParkinsonDoloAún no hay calificaciones
- Pentosas FosfatoDocumento39 páginasPentosas FosfatoKarolAún no hay calificaciones
- Lab Bioquimica - AminoacidosDocumento1 páginaLab Bioquimica - Aminoacidosdeosx67% (3)
- 6 GluconeogénesisDocumento15 páginas6 Gluconeogénesisdora100% (1)
- ProteinasDocumento5 páginasProteinasRené Cariño100% (2)
- PROTEINASDocumento26 páginasPROTEINASMariselCielo100% (1)
- El Ciclo de KrebsDocumento16 páginasEl Ciclo de KrebsClemente Peralta Serrano100% (2)
- SUEROSDocumento2 páginasSUEROSValeska FigueroaAún no hay calificaciones
- Bioquimica Del Agua y Tampones FisiologicosDocumento24 páginasBioquimica Del Agua y Tampones FisiologicospedrojacintohAún no hay calificaciones
- Las ProteinasDocumento14 páginasLas Proteinasanabely chumacero lizana50% (2)
- El Agua - PPT BioquimicaDocumento14 páginasEl Agua - PPT BioquimicaIsabella BlackAún no hay calificaciones
- Cuestionario Biosintesis de AminoacidosDocumento7 páginasCuestionario Biosintesis de AminoacidosLeandro Torres CalabiAún no hay calificaciones
- Glucólisis, Gluconeogénesis y Ruta de Las Pentosas FosfatoDocumento26 páginasGlucólisis, Gluconeogénesis y Ruta de Las Pentosas FosfatoMarcelo Kim100% (1)
- Metabolismo Del GlucógenoDocumento20 páginasMetabolismo Del Glucógenoqlawdya123Aún no hay calificaciones
- Metabolismo de ProteinasDocumento21 páginasMetabolismo de ProteinasRonald Astucuri PalaciosAún no hay calificaciones
- 2 Clasificación de Los Aminoácidos en Función de La Naturaleza de Su Cadena LateralDocumento4 páginas2 Clasificación de Los Aminoácidos en Función de La Naturaleza de Su Cadena LateralAsier SantamaríaAún no hay calificaciones
- PROTEÍNASDocumento14 páginasPROTEÍNASJose Maravi100% (2)
- Beta-Oxidación de Los Ácidos GrasosDocumento9 páginasBeta-Oxidación de Los Ácidos GrasosJohanna CortésAún no hay calificaciones
- LipidosDocumento5 páginasLipidosJosé Luis FerrerAún no hay calificaciones
- 24 Metabolismo LípidosDocumento31 páginas24 Metabolismo LípidosClau VargasAún no hay calificaciones
- Nucleósidos y NucleótidosDocumento2 páginasNucleósidos y NucleótidosAdriana RodriguezAún no hay calificaciones
- Introduccion Al MetabolismoDocumento33 páginasIntroduccion Al MetabolismoJosé RafaelAún no hay calificaciones
- Vía de Las Pentosas FosfatoDocumento9 páginasVía de Las Pentosas FosfatoJeff Willian Rosas QuinaAún no hay calificaciones
- TRIPSINADocumento1 páginaTRIPSINAJorge HueichaAún no hay calificaciones
- wuolah-free-PREGUNTAS IMPORTANTES PRIMER PARCIALDocumento37 páginaswuolah-free-PREGUNTAS IMPORTANTES PRIMER PARCIALCESAR SALDANA RAMIREZAún no hay calificaciones
- Manipulacion GeneticaDocumento2 páginasManipulacion GeneticaÁngel HernandezAún no hay calificaciones
- Aislamiento de Glucogeno Del Higado de PolloDocumento7 páginasAislamiento de Glucogeno Del Higado de PolloGise HinostrozaAún no hay calificaciones
- Lo de VirologíaDocumento18 páginasLo de VirologíaFernando RiañosAún no hay calificaciones
- Clase 5. Componentes Mayoritarios en Alimentos UplaDocumento12 páginasClase 5. Componentes Mayoritarios en Alimentos UplaarkangeljsmAún no hay calificaciones
- Bio 2 33Documento2 páginasBio 2 33María GtAún no hay calificaciones
- Caso Clinico Farmacologia TeoriaDocumento14 páginasCaso Clinico Farmacologia TeoriaSley Victoria Mavila JimenesAún no hay calificaciones
- Bioenergética Repaso - Lucas Grabiel Liotta - UCU 2022Documento14 páginasBioenergética Repaso - Lucas Grabiel Liotta - UCU 2022Leandro VillalbaAún no hay calificaciones
- S2 PPT Biología General 1Documento25 páginasS2 PPT Biología General 1Milagros ContrerasAún no hay calificaciones
- Actividad 2Documento6 páginasActividad 2Lina Pedraza GaonaAún no hay calificaciones
- Informe Biologia EnzimasDocumento12 páginasInforme Biologia EnzimasMarq TintaAún no hay calificaciones
- FundetecDocumento6 páginasFundetecalexandra leonAún no hay calificaciones
- Examen Unidad III de Bioquimica GeneralDocumento4 páginasExamen Unidad III de Bioquimica GeneralDiego ChecaAún no hay calificaciones
- Organización Cuerpo Humano TallerDocumento3 páginasOrganización Cuerpo Humano TallerEdwin RodriguezAún no hay calificaciones
- Aparato o Complejo de GolgiDocumento13 páginasAparato o Complejo de GolgiNicolas HerreraAún no hay calificaciones
- Mapa Membrana PlasmaticaDocumento1 páginaMapa Membrana Plasmaticavictor reaAún no hay calificaciones
- Resume NDocumento5 páginasResume NJordan Josue TorresAún no hay calificaciones
- Cuerpos NuclearesDocumento4 páginasCuerpos NucleareszaraAún no hay calificaciones
- Adherencia Microbiana I - II (N°4-5)Documento8 páginasAdherencia Microbiana I - II (N°4-5)LESLIE JEANETTE MELGAR LOPEZAún no hay calificaciones
- Lipidos TeoriaDocumento42 páginasLipidos TeoriaBrayerAún no hay calificaciones
- Celula Procariota Eucariota Estructura OrganelosDocumento6 páginasCelula Procariota Eucariota Estructura OrganelosLaura Perez0% (1)
- Lectura 2 - 4º Grado Cyt - III Unidad - Docx-1Documento3 páginasLectura 2 - 4º Grado Cyt - III Unidad - Docx-1Cecilia Sairitupac PachecoAún no hay calificaciones
- Informe N°04 - Proteínas - IDocumento8 páginasInforme N°04 - Proteínas - IDenisse Aracely Usecca CondoriAún no hay calificaciones
- AUTOTROFOSokDocumento42 páginasAUTOTROFOSokYudith Rosaluz Imata CondoriAún no hay calificaciones
- Bioenergética y Tipos de Reacciones BioquímicasDocumento49 páginasBioenergética y Tipos de Reacciones BioquímicasRocio LópezAún no hay calificaciones
- Tema 3.farmacodinamiaDocumento43 páginasTema 3.farmacodinamiaClara lisbeth Segundo romeroAún no hay calificaciones
- Biofísica y BioquímicaDocumento14 páginasBiofísica y BioquímicamacckofAún no hay calificaciones