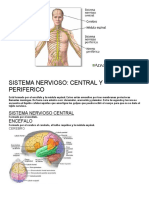
También podría gustarte
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Desarrollo Ontogenético Del Sistema NerviosoDocumento24 páginasDesarrollo Ontogenético Del Sistema NerviosoRadhamePerez100% (7)
- Guía de Entrevista Clínica para Adultos - Manuel MuñozDocumento3 páginasGuía de Entrevista Clínica para Adultos - Manuel MuñozNathaly Berrío100% (1)
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- El nervio periférico: Estructura y funciónDe EverandEl nervio periférico: Estructura y funciónAún no hay calificaciones
- Desarrollo Embrionario Del Sistema Nervioso Central y Órganos de Los Sentidos - RevisiónDocumento9 páginasDesarrollo Embrionario Del Sistema Nervioso Central y Órganos de Los Sentidos - RevisiónCristhiam Anderson B WAún no hay calificaciones
- Neuropsicología Del Desarrollo Infantil 2Documento5 páginasNeuropsicología Del Desarrollo Infantil 2Cristian Wollfan TrujilloAún no hay calificaciones
- Desarrollo Intrauterino de Los Órganos de Los SentidosDocumento9 páginasDesarrollo Intrauterino de Los Órganos de Los SentidosCarlos Zanns CarpioAún no hay calificaciones
- Sistema NerviosoDocumento24 páginasSistema NerviosoAngeliannis CardenadAún no hay calificaciones
- Desarrollo Del Sistema Nervioso CentralDocumento24 páginasDesarrollo Del Sistema Nervioso CentralJoan Alexander GomezAún no hay calificaciones
- Sistema NerviosoDocumento13 páginasSistema NerviosoGarzain ReyesAún no hay calificaciones
- Morfofisiologia Humana II Conferencia 01Documento12 páginasMorfofisiologia Humana II Conferencia 01Jose CadenasAún no hay calificaciones
- NEURULACIÓN Juev. 10 de Dic.Documento13 páginasNEURULACIÓN Juev. 10 de Dic.pablo cardonaAún no hay calificaciones
- Desarrollo Ontogenetico Del Sistema NerviosoDocumento11 páginasDesarrollo Ontogenetico Del Sistema NerviosoEimy PerezAún no hay calificaciones
- Valencia Lagunas Yuridia 3pm4 EmbriologíaDocumento5 páginasValencia Lagunas Yuridia 3pm4 EmbriologíaYuri ValenciaAún no hay calificaciones
- 3 Carpeta de EmbriologíaDocumento129 páginas3 Carpeta de Embriologíaemelin garciaAún no hay calificaciones
- Para Saber Más 2Documento20 páginasPara Saber Más 2belen solisAún no hay calificaciones
- Ao-1 MFH IiDocumento16 páginasAo-1 MFH Iiyosman lópezAún no hay calificaciones
- Embriología Del Sistema Nervioso Vesículas Primarias y SecundariasDocumento7 páginasEmbriología Del Sistema Nervioso Vesículas Primarias y SecundariasXavier GuevaraAún no hay calificaciones
- Fases Del DesarrolloDocumento32 páginasFases Del DesarrollomariaAún no hay calificaciones
- Erickson Aarón Martínez Tarea 6 HuehueDocumento10 páginasErickson Aarón Martínez Tarea 6 HuehueAndrea VillatoroAún no hay calificaciones
- Tema 9Documento14 páginasTema 9arancharr01Aún no hay calificaciones
- 2lectura - Desarrollo Morfológico Del SNDocumento6 páginas2lectura - Desarrollo Morfológico Del SNLilian Daniela Añamuro DíazAún no hay calificaciones
- Desarrollo CognitivoDocumento10 páginasDesarrollo CognitivoArturo Said Langarica TapiaAún no hay calificaciones
- Morfofisiologia Humana 2 Actividad OrientadoraDocumento12 páginasMorfofisiologia Humana 2 Actividad Orientadoramaria torres100% (1)
- Tubo NeuralDocumento3 páginasTubo NeuralDante Huaman CamposAún no hay calificaciones
- Resumen Del Sistema NerviosoDocumento5 páginasResumen Del Sistema NerviosoAtenea 03Aún no hay calificaciones
- 1.neuropsicologia Del Desarrollo Somatosensorial y MotorDocumento17 páginas1.neuropsicologia Del Desarrollo Somatosensorial y MotorNohemy CastañoAún no hay calificaciones
- NEUROCIENCIASDocumento17 páginasNEUROCIENCIASTohru UzumakiAún no hay calificaciones
- Cedva 9Documento22 páginasCedva 9Andrea JscAún no hay calificaciones
- Guia de Embrio Intensa para Muchachos IntensosDocumento56 páginasGuia de Embrio Intensa para Muchachos IntensosAna Amy Aguilar MartinezAún no hay calificaciones
- Universidad para El Desarrollo Andino Facultad Humanidades Y Ciencias SocialesDocumento43 páginasUniversidad para El Desarrollo Andino Facultad Humanidades Y Ciencias SocialesJhakilin m Huacho JanampaAún no hay calificaciones
- Embriologia Cuestionario Unidad Sistema Nervioso Origen EmbriologicoDocumento8 páginasEmbriologia Cuestionario Unidad Sistema Nervioso Origen EmbriologicoSebastian PerezAún no hay calificaciones
- EMBRIOLOGIA Conferencia 1a.10!02!2018Documento17 páginasEMBRIOLOGIA Conferencia 1a.10!02!2018Nury CruzAún no hay calificaciones
- Desarrollo Ontogenetico Del SNDocumento14 páginasDesarrollo Ontogenetico Del SNStephany Marte100% (3)
- Trabajo #1 EmbriologiaDocumento32 páginasTrabajo #1 EmbriologiaJennifer Katherine Guzman GutierrezAún no hay calificaciones
- Lectura Complementaria. Desarrollo Embrionario. Organogénesis.Documento25 páginasLectura Complementaria. Desarrollo Embrionario. Organogénesis.gabrielaAún no hay calificaciones
- Desarrollo Ontogenito Delsistema Nervioso RafaelDocumento16 páginasDesarrollo Ontogenito Delsistema Nervioso RafaelOdalis De JesúsAún no hay calificaciones
- Fases Del Proceso de Desarrollo Del Sistema Nervioso en Él Infante PDFDocumento7 páginasFases Del Proceso de Desarrollo Del Sistema Nervioso en Él Infante PDFMision Juvenil Ipuc ParnasoAún no hay calificaciones
- Embriologia y NeurodesarrolloDocumento4 páginasEmbriologia y NeurodesarrolloMoises del CidAún no hay calificaciones
- Desarrollo Embrionario 1Documento24 páginasDesarrollo Embrionario 1Pedro Manuel Fajardo YanacAún no hay calificaciones
- Foro 14Documento5 páginasForo 14Nikole HernándezAún no hay calificaciones
- Bases Del Neurodesarrollo PDFDocumento182 páginasBases Del Neurodesarrollo PDFgemma galvezAún no hay calificaciones
- Diapositiva de Embriogenesis Del Sistema NerviosoDocumento62 páginasDiapositiva de Embriogenesis Del Sistema NerviosoIvo AntonyAún no hay calificaciones
- Tema1 AO123Documento16 páginasTema1 AO123MARLYN MATAAún no hay calificaciones
- Clase 3 EmbriologiaDocumento32 páginasClase 3 EmbriologiaMarcial Hernández MercadoAún no hay calificaciones
- SNC EmbriologíaDocumento16 páginasSNC EmbriologíaFanny Sarahí Pérez SánchezAún no hay calificaciones
- Resumen Clase 2Documento12 páginasResumen Clase 2Pablo GarciaAún no hay calificaciones
- Psicologia Sistema NberviosoDocumento6 páginasPsicologia Sistema NberviosoLupita RamirezAún no hay calificaciones
- Sistema NerviosoDocumento18 páginasSistema NerviosoantonioAún no hay calificaciones
- Informe Desarrollo de Las Capas Embrionarias y Tubo Neural - Semana2Documento12 páginasInforme Desarrollo de Las Capas Embrionarias y Tubo Neural - Semana2Laura MarAún no hay calificaciones
- Proyecto Regeneración de Las Células Nerviosas en AdultosDocumento16 páginasProyecto Regeneración de Las Células Nerviosas en AdultosCONSTANZA VALENTINA BASTIAS TAPIAAún no hay calificaciones
- Embriologia de SNC (Tema 3) - 075253Documento9 páginasEmbriologia de SNC (Tema 3) - 075253Oriana Segnini RamirezAún no hay calificaciones
- Informe NeurocienciasDocumento8 páginasInforme NeurocienciasCésar AntonioAún no hay calificaciones
- 13 Na 1Documento22 páginas13 Na 1Sanatú PsicoterapiaAún no hay calificaciones
- Gould, D. (2015) - Neuroanatomía (5th Edition Ed.) y Purves de NeurocienciaDocumento5 páginasGould, D. (2015) - Neuroanatomía (5th Edition Ed.) y Purves de NeurocienciaFelipe GuzmanAún no hay calificaciones
- Ontogénico Del Sistema NerviosoDocumento18 páginasOntogénico Del Sistema NerviosojonnyAún no hay calificaciones
- Practica Sobre El Desarrollo Del SN para Llenar y Entregar en ClaseDocumento2 páginasPractica Sobre El Desarrollo Del SN para Llenar y Entregar en ClaseMarielle LuloAún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- Presentacion Encuesta Nacional Salud Mental 2015 PDFDocumento22 páginasPresentacion Encuesta Nacional Salud Mental 2015 PDFNathaly BerríoAún no hay calificaciones
- Boletín de Salud Mental DepresiónDocumento16 páginasBoletín de Salud Mental DepresiónNathaly BerríoAún no hay calificaciones
- Técnicas Análisis Diseño Caso 1Documento14 páginasTécnicas Análisis Diseño Caso 1Nathaly BerríoAún no hay calificaciones
- Escala de Sociotropía-AutonomíaDocumento15 páginasEscala de Sociotropía-AutonomíaNathaly BerríoAún no hay calificaciones
- 5 - ATIENZA M, Cápitulo I (Bioética y Argumentación)Documento31 páginas5 - ATIENZA M, Cápitulo I (Bioética y Argumentación)Alfonsina Del SurAún no hay calificaciones
- Naturaleza Jurídica de Las Declaraciones InternacionalesDocumento11 páginasNaturaleza Jurídica de Las Declaraciones InternacionalesNathaly BerríoAún no hay calificaciones
- Infantofilia Pedofilia y Hebefilia-RAFAEL GARCÍADocumento20 páginasInfantofilia Pedofilia y Hebefilia-RAFAEL GARCÍANathaly BerríoAún no hay calificaciones
- Adultos Con Trastornos de AprendizajeDocumento11 páginasAdultos Con Trastornos de AprendizajeNathaly BerríoAún no hay calificaciones
- El CardadoDocumento4 páginasEl CardadoOlger Aragon BerlangaAún no hay calificaciones
- COMA (Guía Completa)Documento10 páginasCOMA (Guía Completa)G'Belen SánchezAún no hay calificaciones
- NEUROIMAGENESDocumento2 páginasNEUROIMAGENESmaria lizeth gutierrez vargasAún no hay calificaciones
- AutismoDocumento9 páginasAutismoAdrian Guexpal TepoxAún no hay calificaciones
- Tesis 64257Documento67 páginasTesis 64257Dayelis PaezAún no hay calificaciones
- Neurogénesis Del MovimientoDocumento10 páginasNeurogénesis Del MovimientoMaria Laura Vega MillicanAún no hay calificaciones
- Partes Del Cerebro Humano y FuncionesDocumento6 páginasPartes Del Cerebro Humano y FuncionesVergel L. Dayana PaolaAún no hay calificaciones
- c11 CerebeloDocumento50 páginasc11 CerebeloMiguelMonsefúAún no hay calificaciones
- 1ro Psicologia 2do TrimDocumento10 páginas1ro Psicologia 2do TrimAlvaro Lapaca SupoAún no hay calificaciones
- EXAMEN DE SIMULACRO FUERTES II 29 de MayoDocumento4 páginasEXAMEN DE SIMULACRO FUERTES II 29 de MayoShieda OwO?Aún no hay calificaciones
- Dossier de Integracion Sensorial Y TEA para Profesionales y Familias PDFDocumento63 páginasDossier de Integracion Sensorial Y TEA para Profesionales y Familias PDFJesus Fajardo100% (1)
- Sistema Nervioso Central y PerifericoDocumento3 páginasSistema Nervioso Central y PerifericoAndres Quiroz100% (1)
- Síndromes CerebelososDocumento29 páginasSíndromes CerebelososPRACTICA MEDICA IIAún no hay calificaciones
- 2022 CerebeloDocumento16 páginas2022 CerebeloDenisse Nickole SemperteguiAún no hay calificaciones
- Sistema Nervioso CentralDocumento25 páginasSistema Nervioso CentralMargarita FloresAún no hay calificaciones
- Tejido Nervioso, Sistema Nervioso Actual Enero 2018Documento142 páginasTejido Nervioso, Sistema Nervioso Actual Enero 2018Wendy Guissella TaipeAún no hay calificaciones
- Original Guia Teorica Ciencias Naturalez 2año I Lapso DefinitivaDocumento7 páginasOriginal Guia Teorica Ciencias Naturalez 2año I Lapso DefinitivaJosW12Aún no hay calificaciones
- El Sistema Nervioso CentralDocumento12 páginasEl Sistema Nervioso CentralJamer MartosAún no hay calificaciones
- Resumen Neuro Fisiologia Ibjg¡Documento18 páginasResumen Neuro Fisiologia Ibjg¡italo4bryan4jurado4gAún no hay calificaciones
- El Cerebro 200307 Downloable 1193026Documento9 páginasEl Cerebro 200307 Downloable 1193026Gloria Magali BlasichAún no hay calificaciones
- La Formación Reticular Y El Sistema Límbico: Capítulo 9Documento16 páginasLa Formación Reticular Y El Sistema Límbico: Capítulo 9Gabriela Abella CastellanosAún no hay calificaciones
- Guia FisioterapiaDocumento58 páginasGuia FisioterapiaHarry Marquez Leal100% (1)
- 1.-Formacion - Reticular - 2021Documento95 páginas1.-Formacion - Reticular - 2021Isaac CasillasAún no hay calificaciones
- Portafolio de Biologia Eloiza Arias ZalduaDocumento8 páginasPortafolio de Biologia Eloiza Arias Zalduaeloiza ariasAún no hay calificaciones
- Sesión 1 Tema 1 y 2 Parte I 03MNEUDocumento66 páginasSesión 1 Tema 1 y 2 Parte I 03MNEULauraSánchezGiraldoAún no hay calificaciones
- Practica de Laboratorio Observo El Cerebro de Un VertebradoDocumento3 páginasPractica de Laboratorio Observo El Cerebro de Un VertebradoDiego MontañaAún no hay calificaciones
- Grandes Síndromes Toxicológicos. Cuadros AnatomopatológicosDocumento23 páginasGrandes Síndromes Toxicológicos. Cuadros Anatomopatológicosmelani quispeAún no hay calificaciones
- Sistema VestibularDocumento22 páginasSistema VestibularCintia100% (1)
- Yacila-Cunya-Abish-Esther-Guia 2Documento9 páginasYacila-Cunya-Abish-Esther-Guia 2Abish Yacila CunyaAún no hay calificaciones
- NEUROPATOLOGIA-PARTE 1 y 2Documento20 páginasNEUROPATOLOGIA-PARTE 1 y 2María Alvarado PachasAún no hay calificaciones