También podría gustarte
- Conservacion-Extraccion Leche MaternaDocumento22 páginasConservacion-Extraccion Leche MaternaCaro VargasAún no hay calificaciones
- Camilla-Mora-Despertar A Tu Lado PDFDocumento191 páginasCamilla-Mora-Despertar A Tu Lado PDFcamilaAún no hay calificaciones
- Lactancia MaternaDocumento46 páginasLactancia MaternaNanci A JiménezAún no hay calificaciones
- Revista Lactancia MaternaDocumento16 páginasRevista Lactancia MaternaRomer Juanez AramayoAún no hay calificaciones
- Lactancia Materna en PrematurosDocumento18 páginasLactancia Materna en PrematurosCamila SiebzehnerAún no hay calificaciones
- Impacto de La Lactancia en El Crecimiento de Los Bebes PDFDocumento33 páginasImpacto de La Lactancia en El Crecimiento de Los Bebes PDFNatalia TorricoAún no hay calificaciones
- La Zorra y Las UvasDocumento15 páginasLa Zorra y Las UvasMayte CrespinAún no hay calificaciones
- Caperucita RojaDocumento48 páginasCaperucita RojaLucila SantelAún no hay calificaciones
- Procesos Biofisiológicos en La LM de La Mama PDFDocumento23 páginasProcesos Biofisiológicos en La LM de La Mama PDFLeonardo GonzalezAún no hay calificaciones
- Sucedáneos de La Leche Materna y Otras FormulasDocumento5 páginasSucedáneos de La Leche Materna y Otras FormulasLtMorelosAún no hay calificaciones
- Embarazo Añoso y Adolecente MonografiaDocumento23 páginasEmbarazo Añoso y Adolecente MonografiaSILVA100% (1)
- Ancestria EcuacionesDocumento51 páginasAncestria EcuacionesDonajirum PSAún no hay calificaciones
- Fisiologia de La LactanciaDocumento18 páginasFisiologia de La LactanciaPolette Cartes GaticaAún no hay calificaciones
- Signo LinguísticoDocumento61 páginasSigno LinguísticoJoan Paolo Gonzáles Arellano100% (1)
- Lactancia Materna Vs Lactancia ArtificialDocumento24 páginasLactancia Materna Vs Lactancia ArtificialCorina Bizama AravenaAún no hay calificaciones
- Hoja de Calificacion de Maria MelgarDocumento2 páginasHoja de Calificacion de Maria MelgarMilagros Roxana Tito CandiaAún no hay calificaciones
- Práctica de La Odontología Durante El RenacimientoDocumento8 páginasPráctica de La Odontología Durante El RenacimientoMarvin Zapata Zapata100% (1)
- Guia Seres Vertebrados e InvertebradosDocumento6 páginasGuia Seres Vertebrados e InvertebradosLois Valeria Tapia Rivera90% (10)
- Lactancia Materna y Alim ComplementariaDocumento21 páginasLactancia Materna y Alim ComplementariaMicol EscaparlendaAún no hay calificaciones
- Lactancia Materna - Madre Con Enfermedad Tratada Con MedicamentosDocumento37 páginasLactancia Materna - Madre Con Enfermedad Tratada Con MedicamentosJessica Ramírez Rodríguez100% (1)
- Prueba Ingo y DragoDocumento10 páginasPrueba Ingo y DragoMaryMarimán100% (1)
- El Sistema Inmune Del Recién NacidoDocumento2 páginasEl Sistema Inmune Del Recién NacidoJuan Daniel RodríguezAún no hay calificaciones
- Prueba de Ciencias Naturales - 6° BasicoDocumento7 páginasPrueba de Ciencias Naturales - 6° BasicoAnonymous 5WjcH8OVwAún no hay calificaciones
- Nutricion EnteralDocumento14 páginasNutricion EnteralJenniffer Tituana MoranAún no hay calificaciones
- Lactancia Materna en Situaciones EspecialesDocumento43 páginasLactancia Materna en Situaciones EspecialesbrigittedenisseAún no hay calificaciones
- Importancia EmbriologiaDocumento26 páginasImportancia EmbriologiaRuth RodriguezAún no hay calificaciones
- Prueba de "Primer Semestre 2º Básico.: Ciencias NaturalesDocumento4 páginasPrueba de "Primer Semestre 2º Básico.: Ciencias Naturaleslilian cervellon100% (2)
- 08 Diseños Estudios EpidemiologicosDocumento53 páginas08 Diseños Estudios Epidemiologicosapi-2699092071% (7)
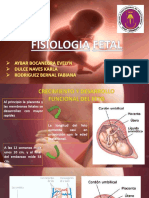
- Fisiologia Fetal y LactanciaDocumento39 páginasFisiologia Fetal y LactanciaSAIDA ARELI RUIZ ACUÑAAún no hay calificaciones
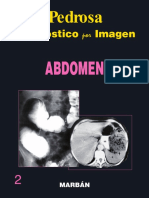
- Pedrosa Abdomen PDFDocumento10 páginasPedrosa Abdomen PDFgildajazmin40% (5)
- Psicologia MedicaDocumento293 páginasPsicologia Medicaapi-370722875% (8)
- Farmacos en La LactanciaDocumento17 páginasFarmacos en La LactanciaCarolinaEspinozaRojasAún no hay calificaciones
- Lactancia MaternaDocumento1 páginaLactancia MaternaFatima YeeAún no hay calificaciones
- Técnicas de Lactancia MaternaDocumento6 páginasTécnicas de Lactancia MaternaAldair Asencios100% (1)
- Composición Nutricional de La Leche Materna y La Fórmula para Prematuros para El Lactante Prematuro - UpToDateDocumento20 páginasComposición Nutricional de La Leche Materna y La Fórmula para Prematuros para El Lactante Prematuro - UpToDateAna Valeria Guerrero VarticovskyAún no hay calificaciones
- Cómo Se Produce La Leche MaternaDocumento3 páginasCómo Se Produce La Leche MaternaLaura Vanessa Sarabia MoralesAún no hay calificaciones
- Nutricion y LactanciaDocumento40 páginasNutricion y Lactanciaapi-3697362100% (1)
- Preparación y LactanciaDocumento6 páginasPreparación y LactanciaAbelardo Diaz VasquezAún no hay calificaciones
- Ventajas y Desventajas de La Lactancia MaternaDocumento10 páginasVentajas y Desventajas de La Lactancia MaternaMeli BlemAún no hay calificaciones
- Triptico Sensibilizacion Lactancia Materna 2 EdicionDocumento2 páginasTriptico Sensibilizacion Lactancia Materna 2 EdicionAndres SuasnavasAún no hay calificaciones
- La Placenta InformeDocumento4 páginasLa Placenta Informekatherinfrias100% (1)
- Anemia Del Recien NacidoDocumento8 páginasAnemia Del Recien NacidoMarleny HCAún no hay calificaciones
- Fortificacion Leche MaternaDocumento1 páginaFortificacion Leche MaternaclaudiaAún no hay calificaciones
- 2.lactancia Materna Anatomia y Fisiologia PDFDocumento21 páginas2.lactancia Materna Anatomia y Fisiologia PDFBailón Hernandez100% (2)
- Lactancia MaternaDocumento3 páginasLactancia MaternaJhonatan RengifoAún no hay calificaciones
- Embarazo MúltipleDocumento27 páginasEmbarazo MúltipleMayra Elizabeth Quiroz FloriánAún no hay calificaciones
- Embarazo y Lactancia Materna Cambios FisiologicosDocumento19 páginasEmbarazo y Lactancia Materna Cambios FisiologicosDaniel TorresAún no hay calificaciones
- Rotafolio Lactancia - 2021Documento10 páginasRotafolio Lactancia - 2021Carol Pinaya AyalaAún no hay calificaciones
- Leche Materna - Leche de VacaDocumento13 páginasLeche Materna - Leche de VacalyliattAún no hay calificaciones
- Apego Inmediato Del Recien NacidoDocumento2 páginasApego Inmediato Del Recien NacidoLeonardoQuero0% (1)
- Triptico LactanciaDocumento2 páginasTriptico LactanciaYaquelin MelendezAún no hay calificaciones
- Crecimiento y Desarrollo Del Recien NacidoDocumento27 páginasCrecimiento y Desarrollo Del Recien NacidoMiguel Ruiz Cobian100% (1)
- 4 Cambios Fisiologicos en El EmbarazoDocumento61 páginas4 Cambios Fisiologicos en El EmbarazoPabloYefersonEspinozaAyoroaAún no hay calificaciones
- Fisiología y Bioquímica Del Parto-BRLDocumento30 páginasFisiología y Bioquímica Del Parto-BRLBeindyAún no hay calificaciones
- Desarollo FetalDocumento20 páginasDesarollo FetalGuadalupe del Carmen González BarradasAún no hay calificaciones
- Nutricion Megalabs Moodulo 1Documento44 páginasNutricion Megalabs Moodulo 1Danilo SantillánAún no hay calificaciones
- Escalas de Valoración Recien NacidoDocumento56 páginasEscalas de Valoración Recien NacidoDIANA LAURA TRINIDAD JIMENEZAún no hay calificaciones
- Estudio de La Lactancia MaternaDocumento35 páginasEstudio de La Lactancia Maternapamarchez100% (1)
- Cambios Fisiologicos Del PuerperioDocumento11 páginasCambios Fisiologicos Del PuerperioKAREN ORTIZAún no hay calificaciones
- Maniobras de LeopoldDocumento26 páginasManiobras de LeopoldZPol RodriAún no hay calificaciones
- Historia Lactancia MaternaDocumento10 páginasHistoria Lactancia Maternafrancisco morales100% (1)
- Ictericia X Lactancia MDocumento6 páginasIctericia X Lactancia MAnaly Cajusol DiazAún no hay calificaciones
- El Reflejo de SucciónDocumento1 páginaEl Reflejo de Succiónmonik201989Aún no hay calificaciones
- Respiracion en El PartoDocumento2 páginasRespiracion en El PartokelyAún no hay calificaciones
- Hipotálamo LaCTANCIADocumento6 páginasHipotálamo LaCTANCIAMaria Andreina DiazAún no hay calificaciones
- Protocolo LATCHDocumento29 páginasProtocolo LATCHAna LyaAún no hay calificaciones
- Lactancia Materna - Anatomia y Fisiologia de La MamaDocumento4 páginasLactancia Materna - Anatomia y Fisiologia de La MamabloodyroardAún no hay calificaciones
- Libro LactanciaDocumento75 páginasLibro LactanciaCarolina Herrera Hincapie100% (1)
- Signos de Alarma en El EmbarazoDocumento11 páginasSignos de Alarma en El Embarazoamoalcristal969Aún no hay calificaciones
- Fisiologia de La Glandula Mamaria y LactanciaDocumento9 páginasFisiologia de La Glandula Mamaria y LactanciaRebeca VallesAún no hay calificaciones
- Proyecto 3ero Ciencias B Glangulas Mamaras Aguilar-HonoresDocumento9 páginasProyecto 3ero Ciencias B Glangulas Mamaras Aguilar-HonoresMOOM SublimacionesAún no hay calificaciones
- Lactancia MaternaDocumento15 páginasLactancia Maternamarcel vicencioAún no hay calificaciones
- Lacto GenesisDocumento4 páginasLacto GenesisFabián BalboaAún no hay calificaciones
- Código de Ética y Deontología Médica Del PerúDocumento19 páginasCódigo de Ética y Deontología Médica Del Perúapi-370007550% (2)
- GuÍa Practicas Usmp 2008 10 Marzo 2008Documento54 páginasGuÍa Practicas Usmp 2008 10 Marzo 2008api-3700075Aún no hay calificaciones
- Clase 9 - Estudios Cos EpidemiologicosDocumento70 páginasClase 9 - Estudios Cos Epidemiologicosapi-3711483100% (1)
- Guia de Recursos Web en SaludDocumento124 páginasGuia de Recursos Web en Saludapi-3711609Aún no hay calificaciones
- Lactancia Materna - Anatomia y Fisiologia de La MamaDocumento4 páginasLactancia Materna - Anatomia y Fisiologia de La MamabloodyroardAún no hay calificaciones
- Absorción y Secreción de Electrolitos y AguaDocumento1 páginaAbsorción y Secreción de Electrolitos y Aguaapi-370007567% (6)
- El Cerebro Emocional PDFDocumento9 páginasEl Cerebro Emocional PDFMiguel RamosAún no hay calificaciones
- 2primeros Auxilios en Urgencias y Emergencias MédicasDocumento228 páginas2primeros Auxilios en Urgencias y Emergencias MédicasClansy TurkAún no hay calificaciones
- 2dc184e25de7e79bbd64a9474db81a456936f1d1f758c4263cc411558723386fDocumento55 páginas2dc184e25de7e79bbd64a9474db81a456936f1d1f758c4263cc411558723386fEdilson Alcides Vargas AroneAún no hay calificaciones
- Clases Diversas de Monos y Sus NombresDocumento6 páginasClases Diversas de Monos y Sus Nombressalomon centeno maguiñaAún no hay calificaciones
- Giuseppe ArcimboldoDocumento3 páginasGiuseppe Arcimboldoandoba2013Aún no hay calificaciones
- Tronco EncefálicoDocumento15 páginasTronco EncefálicoJeraldyne Stefani Lucas AguirreAún no hay calificaciones
- Historia de La Hoteleria en TacnaDocumento13 páginasHistoria de La Hoteleria en TacnaVhiirGhiiniia DheysiiAún no hay calificaciones
- Lo Importànte en El CantoDocumento2 páginasLo Importànte en El CantoNickO.LateAún no hay calificaciones
- Trabajo Práctico #2 Alfabetización-1Documento19 páginasTrabajo Práctico #2 Alfabetización-1NoeliaAún no hay calificaciones
- Zoologicos Humanos PDFDocumento3 páginasZoologicos Humanos PDFBeto PanquequeAún no hay calificaciones
- Informe Biologia.. S. Digestivo, Circulatorio, RespiratorioDocumento29 páginasInforme Biologia.. S. Digestivo, Circulatorio, RespiratorioMaickol F BalarezoAún no hay calificaciones
- Vale Diccionario MapucheDocumento4 páginasVale Diccionario MapucheKatherin Angelica MendezAún no hay calificaciones
- Las Momias ChinchorroDocumento3 páginasLas Momias ChinchorroPablo Henríquez CarrascoAún no hay calificaciones
- NutricionDocumento9 páginasNutricionCarlos Eladio Sanchez RoselAún no hay calificaciones
- CitricosDocumento182 páginasCitricosvictoria ariasAún no hay calificaciones
- Taller Lectura ExploracionesDocumento2 páginasTaller Lectura Exploracionessocialcomfenalco50% (2)
- Biomecanica Evaluacion PosturalDocumento5 páginasBiomecanica Evaluacion PosturalAnonymous sR39eVAún no hay calificaciones
- Camelidos SudamericanosDocumento25 páginasCamelidos SudamericanosFrancisco Adriano ChilquilloAún no hay calificaciones
- 01 Introduccion A La Fisiología CelularDocumento54 páginas01 Introduccion A La Fisiología CelularRamon Angel Cruz SaldarriagaAún no hay calificaciones