También podría gustarte
- Sistema Digestivo TrabajoDocumento14 páginasSistema Digestivo TrabajoKevin Fox AttoAún no hay calificaciones
- Control de Los Microorganismos Por Agentes Fisicos y Quimicos 2Documento15 páginasControl de Los Microorganismos Por Agentes Fisicos y Quimicos 2Jose Luis BustilloAún no hay calificaciones
- Metabolismo Es El Conjunto de Procesos Físicos y Químicos y de Reacciones A Las Que Está Sujeta Una CélulaDocumento3 páginasMetabolismo Es El Conjunto de Procesos Físicos y Químicos y de Reacciones A Las Que Está Sujeta Una Célulawandres270Aún no hay calificaciones
- DETERMINANTES DE SALUD MilenaDocumento4 páginasDETERMINANTES DE SALUD MilenaMilena ReyesAún no hay calificaciones
- Proyecto de MetodologiaDocumento30 páginasProyecto de MetodologiaMabel Maytee Morocho Quizhpe0% (1)
- Ejercicios de Tipos de PárrafosDocumento4 páginasEjercicios de Tipos de PárrafosIsaul SantamariaAún no hay calificaciones
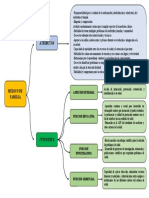
- El Monitoreo EstratégicoDocumento8 páginasEl Monitoreo EstratégicoMaria LealAún no hay calificaciones
- Anaerobiosis o AnaerobismoDocumento1 páginaAnaerobiosis o AnaerobismoJosue David Martines Reyes100% (1)
- Instituciones SocioculturalesDocumento2 páginasInstituciones SocioculturalesCel Seb100% (2)
- Conclusiones de La Calidad de VidaDocumento12 páginasConclusiones de La Calidad de VidaCleveland Chavez Soto100% (1)
- Modelo de Intervencion para La SaludDocumento14 páginasModelo de Intervencion para La SaludAlexis Magaña100% (1)
- Estructura Evolutiva o Psicología Evolutiva (Resumen)Documento16 páginasEstructura Evolutiva o Psicología Evolutiva (Resumen)Claudia Contreras FloresAún no hay calificaciones
- Glosario Terminos HidroterapiaDocumento3 páginasGlosario Terminos Hidroterapiakatherinne GómezAún no hay calificaciones
- CAP 1 - El Comportamiento en Materia de Salud y La Educación Sanitaria (SEM 3)Documento5 páginasCAP 1 - El Comportamiento en Materia de Salud y La Educación Sanitaria (SEM 3)Ivan Solsol ruizAún no hay calificaciones
- Teorias Psicosocial Del EnvejecimientoDocumento1 páginaTeorias Psicosocial Del EnvejecimientoAngela SolisAún no hay calificaciones
- Ensayo motivacion-DESARROLLO HUMANODocumento5 páginasEnsayo motivacion-DESARROLLO HUMANOjanines johana barraza severicheAún no hay calificaciones
- BIOENERGETICADocumento18 páginasBIOENERGETICARossibel100% (1)
- La SaludDocumento22 páginasLa SaludStefaniAún no hay calificaciones
- Ensayo FisioterapiaDocumento6 páginasEnsayo Fisioterapializ fernanda guevara molinaAún no hay calificaciones
- Atencion Primaria Integral en SaludDocumento41 páginasAtencion Primaria Integral en SaludCamiOchoaAún no hay calificaciones
- Antecedentes: 2. Objetivos de La CifDocumento8 páginasAntecedentes: 2. Objetivos de La CifEli QuiMarAún no hay calificaciones
- Características Del Desarrollo Integral Del NiñoDocumento12 páginasCaracterísticas Del Desarrollo Integral Del NiñoMirigan100% (1)
- Columna Vertebral PDFDocumento12 páginasColumna Vertebral PDFAlex HinrichsenAún no hay calificaciones
- Dorothea Orem DiapositivasDocumento9 páginasDorothea Orem DiapositivasLuisa TrampeAún no hay calificaciones
- Informe Final EnfermeriaDocumento51 páginasInforme Final EnfermeriaevelinAún no hay calificaciones
- Tesis de Actividades Recreativas Del Adulto Mayor de TrujilloDocumento21 páginasTesis de Actividades Recreativas Del Adulto Mayor de Trujillopires99100% (2)
- Tema 2. Bases Teóricas de La Educación para La SaludDocumento22 páginasTema 2. Bases Teóricas de La Educación para La SaludZCIAún no hay calificaciones
- Ser Biopsicosocial Trab 1Documento8 páginasSer Biopsicosocial Trab 1Alby Rivera MataAún no hay calificaciones
- Cuidados Integrales para El NiñoDocumento20 páginasCuidados Integrales para El NiñoCarlos Borja ChurampiAún no hay calificaciones
- Salud Comunitaria y FamiliarDocumento28 páginasSalud Comunitaria y FamiliarMarilyn LopezAún no hay calificaciones
- Funciones de Los NucleótidosDocumento3 páginasFunciones de Los Nucleótidosbetincarlos3010Aún no hay calificaciones
- Informe Discapacidad Charla EducativaDocumento16 páginasInforme Discapacidad Charla EducativaAndrea omayri Quispe alconAún no hay calificaciones
- Orientaciones MetodologicasDocumento31 páginasOrientaciones MetodologicasYonherisAún no hay calificaciones
- HidrocefaliaDocumento7 páginasHidrocefaliapaolaAún no hay calificaciones
- Ambiente Terapeutico y Respuestas Humanas BasicasDocumento28 páginasAmbiente Terapeutico y Respuestas Humanas Basicaselvia2321Aún no hay calificaciones
- CAPÍTULO 14 - Desarrollo de Los MúsculosDocumento10 páginasCAPÍTULO 14 - Desarrollo de Los MúsculosKenji DiazAún no hay calificaciones
- Proyecto Cambios Emocionales 2020Documento27 páginasProyecto Cambios Emocionales 2020Camila Vanegas0% (2)
- Reflejos Del Recien Nacido PDFDocumento9 páginasReflejos Del Recien Nacido PDFLisbeth SanchezAún no hay calificaciones
- Atrofia MuscularDocumento8 páginasAtrofia MuscularShely Arriaga100% (1)
- Mapa Mental SISTEMA EDUCATIVO VENEZOLANODocumento6 páginasMapa Mental SISTEMA EDUCATIVO VENEZOLANOWilliam HernandezAún no hay calificaciones
- JesusDocumento30 páginasJesusJesus DiazAún no hay calificaciones
- Funciones Del Medico FamiliarDocumento1 páginaFunciones Del Medico FamiliarTatiana OrtegaAún no hay calificaciones
- Mapa Mental Determinantes de La Salud y La Educacion SanitariaDocumento3 páginasMapa Mental Determinantes de La Salud y La Educacion SanitariaNicolle Van100% (2)
- Materno Infantil 1 - Unidad 1Documento14 páginasMaterno Infantil 1 - Unidad 1Endermary BrachoAún no hay calificaciones
- Influencias Del Maestro Simón Rodriguez en El LibertadorDocumento5 páginasInfluencias Del Maestro Simón Rodriguez en El Libertadorcarlosapr99Aún no hay calificaciones
- PDFDocumento95 páginasPDFElizabeth Hilafaya DuranAún no hay calificaciones
- Epidemiología Nutricional - MGDocumento17 páginasEpidemiología Nutricional - MGRUBITZEL CALLEJAS LEÓN100% (1)
- El Sistema Nervioso Tiene Dos Grandes DivisionesDocumento4 páginasEl Sistema Nervioso Tiene Dos Grandes DivisionesSimulacionJuanH100% (1)
- Necesidades Básicas de Los Nutrientes Que Son Esenciales para El Desarrollo HumanoDocumento71 páginasNecesidades Básicas de Los Nutrientes Que Son Esenciales para El Desarrollo HumanoMaria SaaAún no hay calificaciones
- Generalidades de La Bioquimica y Kinesiologia TEMA 1 17-10-2021Documento11 páginasGeneralidades de La Bioquimica y Kinesiologia TEMA 1 17-10-2021zulyannys castilloAún no hay calificaciones
- Modelos EpidemiologicosDocumento3 páginasModelos Epidemiologicosbrayan pepeAún no hay calificaciones
- VII.01.Transicion Demografica y EpidemiologicaDocumento30 páginasVII.01.Transicion Demografica y EpidemiologicaVanessa Vargas AlzateAún no hay calificaciones
- Control PrenatalDocumento19 páginasControl PrenatalKarin MarinAún no hay calificaciones
- Reforma Salud PDFDocumento83 páginasReforma Salud PDFjaimeAún no hay calificaciones
- Desarrollo Humano DINAMICADocumento8 páginasDesarrollo Humano DINAMICAdaniel ortizAún no hay calificaciones
- Atencion Integral Del Adulto Mayor en Instituciones SocialesDocumento6 páginasAtencion Integral Del Adulto Mayor en Instituciones SocialesElianet RomeroAún no hay calificaciones
- Venezuela La Misión Niño Jesús Protección de La Mujer Embarazada Democracia Participativa Salud en RevoluciónDocumento6 páginasVenezuela La Misión Niño Jesús Protección de La Mujer Embarazada Democracia Participativa Salud en RevoluciónMargarita TorresAún no hay calificaciones
- Derecho A La Salud y A La Vida. (Mapa Conceptual)Documento2 páginasDerecho A La Salud y A La Vida. (Mapa Conceptual)ismeldaAún no hay calificaciones
- Sistema Venoso de La Cintura EscapularDocumento1 páginaSistema Venoso de La Cintura EscapularRosangel Andrea Chirinos De AbreuAún no hay calificaciones
- Rutas MetabólicasDocumento7 páginasRutas MetabólicasWilmer Almonte SalcedoAún no hay calificaciones
- Inseminacion y Material VeterinarioDocumento31 páginasInseminacion y Material VeterinarioJavierAún no hay calificaciones
- Bioquimica y BiotecnologiaDocumento222 páginasBioquimica y BiotecnologiaJavierAún no hay calificaciones
- Lista Precios para Cliente 2Documento8 páginasLista Precios para Cliente 2JavierAún no hay calificaciones
- Henil CimientosDocumento1 páginaHenil CimientosJavierAún no hay calificaciones
- Interacción Analgésica de Meloxicam e Ibuprofeno en DolorDocumento78 páginasInteracción Analgésica de Meloxicam e Ibuprofeno en DolorJavierAún no hay calificaciones
- Lista Precios para Cliente-1Documento3 páginasLista Precios para Cliente-1JavierAún no hay calificaciones
- DETERMINACIÒN DE PREVALENCIA DE FASCIOLA HEPATICA EN BOVINOS EN LOS CAMALES MUNICIPALES DE LAS CIUDADES DE TULCÀN Y SAN GABRIEL Ecuador PDFDocumento122 páginasDETERMINACIÒN DE PREVALENCIA DE FASCIOLA HEPATICA EN BOVINOS EN LOS CAMALES MUNICIPALES DE LAS CIUDADES DE TULCÀN Y SAN GABRIEL Ecuador PDFJavierAún no hay calificaciones
- Informe Laboratorio - Practica 1 2 3Documento26 páginasInforme Laboratorio - Practica 1 2 3greys olarteAún no hay calificaciones
- 8° Guía 1 Célula y Su ClasificaciónDocumento5 páginas8° Guía 1 Célula y Su Clasificacióncesar villa100% (1)
- Reino Protista 01Documento11 páginasReino Protista 01Melvin Leonel Fernandez100% (5)
- Carbohidratos (CHOS)Documento15 páginasCarbohidratos (CHOS)Martin German ValverdeAún no hay calificaciones
- ¡Descubre El Brillo MágicoDocumento2 páginas¡Descubre El Brillo MágicojoseeAún no hay calificaciones
- Temario de Ciencias Naturales Pruebas Ser Bachiller PDFDocumento2 páginasTemario de Ciencias Naturales Pruebas Ser Bachiller PDFJohan FernandezAún no hay calificaciones
- Guia 2 Biologia Noveno PDFDocumento5 páginasGuia 2 Biologia Noveno PDFSandra Patricia Galeano0% (1)
- PROTEÍNASDocumento20 páginasPROTEÍNASRafael Tinco SolórzanoAún no hay calificaciones
- LA ZEOLITA y Agua AlcalinaDocumento20 páginasLA ZEOLITA y Agua AlcalinaformlessFlowerAún no hay calificaciones
- Cartilla5 Segundo Periodo Nutricion Celular PDFDocumento13 páginasCartilla5 Segundo Periodo Nutricion Celular PDFAngie SachicaAún no hay calificaciones
- 5 Dis. Curr. Cs. Naturales 2017Documento58 páginas5 Dis. Curr. Cs. Naturales 2017Flavia VargasAún no hay calificaciones
- Guia TopfDocumento6 páginasGuia TopfPrueba123100% (1)
- ESTRATEGIAS K Y R EcologiaDocumento3 páginasESTRATEGIAS K Y R EcologiaYeyo Rodriguez100% (2)
- Guia C Nat Grado 8° Ciclo #1 20221Documento4 páginasGuia C Nat Grado 8° Ciclo #1 20221I.E. Nicolás GaviriaAún no hay calificaciones
- Langosta AUSTRALIANA PDFDocumento12 páginasLangosta AUSTRALIANA PDFLiza RmAún no hay calificaciones
- Resumen Completo Biologia Uba Xxi 227655 Downloable 2813239Documento31 páginasResumen Completo Biologia Uba Xxi 227655 Downloable 2813239Matias AlvarezAún no hay calificaciones
- Mentalidad de La Edad de BronceDocumento155 páginasMentalidad de La Edad de BroncePato CapurroAún no hay calificaciones
- Ley #5.629Documento18 páginasLey #5.629Carla InsaurraldeAún no hay calificaciones
- Antología Métodos y TécnicasDocumento192 páginasAntología Métodos y TécnicasSEBASTIAN ANAYA BRAVOAún no hay calificaciones
- Guia Maestro BiologiaDocumento16 páginasGuia Maestro BiologiaJocelyn MendozaAún no hay calificaciones
- Organizacion Celular. Base EstructuralDocumento4 páginasOrganizacion Celular. Base EstructuralWILFREDO BAKERAún no hay calificaciones
- Entrega 1 Escenario 4 Laboratorio - BiologíaDocumento10 páginasEntrega 1 Escenario 4 Laboratorio - BiologíaEntregas Académicas IUPGAún no hay calificaciones
- MIGUEL 5ta RevDocumento14 páginasMIGUEL 5ta RevChichoSaltosAún no hay calificaciones
- Cta1 PaDocumento4 páginasCta1 PaFrank Peña EscamiloAún no hay calificaciones
- Vocabulario Gastronomico CientificoDocumento37 páginasVocabulario Gastronomico CientificomacasqwertAún no hay calificaciones
- Guia Respiracion CelularDocumento1 páginaGuia Respiracion CelularGiseth Maria Martinez VillarrealAún no hay calificaciones
- Ciencias 1dfh Bloque 4 La Reproduccion y La Continuidad de La VidaDocumento34 páginasCiencias 1dfh Bloque 4 La Reproduccion y La Continuidad de La VidaSeverino CenicerosAún no hay calificaciones
- Los 4 Errores de HamerDocumento2 páginasLos 4 Errores de HamerSusana PalaciosAún no hay calificaciones
- FEV Clase 3 Claudio Rama (2008)Documento16 páginasFEV Clase 3 Claudio Rama (2008)Alejandra PazAún no hay calificaciones
- TV Semana 11 2 PDFDocumento62 páginasTV Semana 11 2 PDFVALEFREY victor alegreAún no hay calificaciones