0 calificaciones0% encontró este documento útil (0 votos)
17 vistas26 páginasFunciones Superiores 2 Gulag Free
El documento aborda la comunicación intercelular entre neuronas a través de sinapsis, destacando la clasificación de estas en químicas, eléctricas y mixtas. Se describen los neurotransmisores, su síntesis, funciones y tipos, así como los receptores involucrados en la transmisión de señales. Además, se menciona la coexistencia de neurotransmisores en una misma neurona, conocidos como cotransmisores, y se explican los mecanismos de acción de la acetilcolina.
Cargado por
claudia maldonadoDerechos de autor
© © All Rights Reserved
Nos tomamos en serio los derechos de los contenidos. Si sospechas que se trata de tu contenido, reclámalo aquí.
Formatos disponibles
Descarga como PDF, TXT o lee en línea desde Scribd
0 calificaciones0% encontró este documento útil (0 votos)
17 vistas26 páginasFunciones Superiores 2 Gulag Free
El documento aborda la comunicación intercelular entre neuronas a través de sinapsis, destacando la clasificación de estas en químicas, eléctricas y mixtas. Se describen los neurotransmisores, su síntesis, funciones y tipos, así como los receptores involucrados en la transmisión de señales. Además, se menciona la coexistencia de neurotransmisores en una misma neurona, conocidos como cotransmisores, y se explican los mecanismos de acción de la acetilcolina.
Cargado por
claudia maldonadoDerechos de autor
© © All Rights Reserved
Nos tomamos en serio los derechos de los contenidos. Si sospechas que se trata de tu contenido, reclámalo aquí.
Formatos disponibles
Descarga como PDF, TXT o lee en línea desde Scribd
TEMA 34.
PRINCIPALES NEUROTRANSMISORES EN EL SNC
COMUNICACIÓN INTERCELULAR ENTRE NEURONAS. LA SINAPSIS
Las neuronas son un tipo de tejido excitable capaces de responder a un estímulo y
generar que PA (impulsos eléctricos) que se transmiten por todo el axón hasta llegar a
un botón terminal.
Las neuronas son capaces de transmitir ese impulso nervioso a otras neuronas,
desencadenando una respuesta.
Se creía que la información viajaba por el sistema nervioso en una red continua de
fibras. Golgi defendía esta teoría, la teoría reticular.
Ramón y Cajal propuso que el SN estaba formado por una red de unidades fásicas, las
neuronas. Dedujo que debido a los espacios que existían entre las neuronas, estas se
comunicaban por contacto, no por continuidad.
Sherrington atribuyó el termino sinapsis como el punto
específico donde 2 neuronas se comunican entre sí.
CLASIFICACIÓN DE LAS SINAPSIS
Según la estructura de contacto:
- Componente presináptico
- Componente postsináptico.
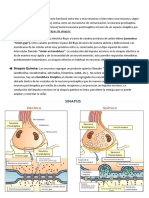
Existen sinapsis químicas, sinapsis eléctricas y sinapsis mixtas.
Sinapsis eléctricas: las membranas presinápticas y las postsinápticas están
prácticamente unidas. La unión se da por medio de unas proteínas (uniones GAP) que
forman un canal que permite el flujo de iones.
Sinapsis química: es la sinapsis más abundante en
vertebrados. Hay una hendidura sináptica donde se
liberan las sustancias químicas o neurotransmisores
almacenados en vesículas desde la neurona presináptica.
La unión neurotransmisor-receptor desencadena cambios
en la permeabilidad de la membrana postsináptica, que
producirá un potencial postsináptico.
Sinapsis mixtas: contiene componentes tanto de sinapsis
químicas o eléctricas.
SINAPSIS QUÍMICA
Es la más abundante en vertebrados. Se clasifica por el tipo de contacto, según el
neurotransmisor que almacena en sus vesículas o según su función.
1. POR EL TIPO DE CONTACTO
Sinapsis neurona-neurona: puede ser:
- Sinapsis axosomáticas: entre axón y el soma. Son señales inhibidoras.
- Sinapsis axodendríticas: entre el axón y las dendritas. Se puede
dar en la rama principal de la dendrita (tallo) o en las espinas
dendríticas, la zona de entrada de señal. Son señales excitadoras.
- Sinapsis axoaxónicas: entre el axón y el axón postsináptico. Son
moduladoras, modulan la cantidad de neurotransmisor que se
liberará a una 3ª neurona.
- Sinapsis dentro-somáticas: entre el soma de una neurona y la
dendrita de la neurona postsináptica.
Sinapsis neurona-efector: el efector puede ser:
- Músculo esquelético.
- Músculo cardíaco.
- Músculo liso.
- Glándula.
2. POR EL TIPO DE NEUROTRANSMISOR
Colinérgicas: se libera Ach.
Aminérgicas: se libera adrenalina, noradrenalina, dopamina o serotonina.
Aminoacidérgicas: glicina, glutamato, ABA, aspartato.
Peptidérgicas: sustancia P, neurotensina, opiáceos (opioides).
Para cada neurotransmisor hay receptores específicos en la membrana de la neurona
postsináptica que por el acoplamiento ligando-receptor desencadena una respuesta.
3. POR FUNCIÓN
Sinapsis excitadora: la unión del neurotransmisor con el receptor genera una
despolarización de la membrana postsináptica.
Sinapsis inhibidora: la unión genera una hiperpolarización (potencial inhibitorio
postsináptico).
Sinapsis moduladora o facilitadora: da lugar a una excitación o una hiperpolarización.
NEUROTRANSMISORES
Biomoléculas que permiten la transmisión de información de una célula nerviosa a otra
mediante la sinapsis.
Características:
- Sintetizados y almacenados en la neurona presináptica. La síntesis se puede
dar tanto en el soma como en la terminación nerviosa.
- Liberados a la hendidura sináptica después de una despolarización de la
membrana de la terminación nerviosa presináptica.
- Una vez liberados, los neurotransmisores deben unirse a receptores
específicos de la membrana pre o postsináptica y dar lugar a una respuesta
rápida y reversible.
- El neurotransmisor se elimina o inactiva en la hendidura sináptica.
Hay algunas excepciones, algunos neurotransmisores como el NO (óxido nítrico) o el
H2S (sulfuro de hidrógeno), que son neurotransmisores gaseosos. Se sintetizan a
demanda de neuronas nitrérgicas o sulfidrérgicas, y se difunden por la membrana del
botón terminal hacia la hendidura sináptica.
Se les clasifica según su estructura:
MOLÉCULA PEQUEÑA
Constituyen un grupo de neurotransmisores heterogéneo.
- Aminas biógenas: dopamina, noradrenalina, adrenalina, serotonina, histamina
y Ach.
- Aminoácidos (aa): glutamato, GABA, glicina.
- Purinas: ATP y adenosina.
MOLÉCULA GRANDE
- Neuropéptidos: endorfinas, encefalinas y sustancia P.
También se clasifica a los neurotransmisores según su función:
NEUROTRANSMISORES EXCITADORES
El acoplamiento de los neurotransmisores con el receptor produce la entrada de calcio
o sodio a la neurona, produciendo la despolarización.
Glutamato: principal neurotransmisor excitador. Presente en el cerebro, en la médula
espinal. Interviene en el 75%.
Otros neurotransmisores excitadores: noradrenalina e histamina.
NEUROTRANSMISORES INHIBIDORES
Pueden modular la excitación, disminuyéndola, ya que abren canales de cloro y
potasio, generando corrientes de entrada de cloro y salida de potasio, generando una
hiperpolarización de la membrana.
GABA: principal neurotransmisor inhibidor. Media tanto la inhibición pre como
postsináptica.
Otros neurotransmisores inhibidores: dopamina, glicina y serotonina.
SÍNTESIS Y METABOLISMO DE LOS NEUROTRANSMISORES
NEUROTRANSMISORES DE MOLÉCULA PEQUEÑA
La síntesis de neurotransmisores (NT) de molécula pequeña se da sobre todo en las
terminaciones nerviosas.
La síntesis de las enzimas necesarias para los neurotransmisores
ocurre en el soma, y luego son transportadas hasta los botones
terminales por el transporte axonal anterógrado.
Las moléculas precursoras para la síntesis de neurotransmisores
son captadas en las terminaciones nerviosas debido a
transportadores de la membrana presináptica.
Los neurotransmisores, una vez sintetizados, son cargados en las
vesículas mediante proteínas transportadoras.
Las vesículas que contienen el neurotransmisor de molécula
pequeña se ven claras en el microscopio, por lo que se denominan vesículas pequeñas
de centro claro, que se desplazan y fusionan con la membrana de la neurona
presináptica por mecanismos dependientes de calcio. Una vez fusionada, se liberan los
neurotransmisores por exocitosis en la hendidura sináptica. El NT se une a los
receptores pre y postsinápticos para ejercer su efecto biológico.
Los NT son eliminados por recaptación o inactivación, a través de enzimas que
recuperan el precursor, que es recaptado por proteínas transportadoras de la
membrana presináptica.
Los NT de molécula pequeña tienen una acción rápida, y generan la mayoría de las
respuestas inmediatas, sensitivas y motoras.
NEUROTRANSMISORES DE MOLÉCULA GRANDE (NEUROPÉPTIDOS)
Los neuropéptidos son sintetizados en el soma de la neurona. Como
cualquier proteína, su síntesis requiere la transcripción de DNA y la
traducción de mRNA. Se sintetizan como prepropéptidos, que sufren un
procesado que les escinde en péptidos más pequeños. Su procesamiento
ocurre en el retículo endoplásmico, donde se forma el propéptido, que
atraviesa el Golgi y se empaqueta en vesículas grandes de núcleo denso.
Estas vesículas son transportadas a los botones terminales por el axón y
a través de los microtúbulos por transporte axonal anterógrado rápido.
Los neuropéptidos se liberan por exocitosis en la hendidura sináptica
por mecanismos dependientes de calcio. Una vez liberados se acoplan a
receptores específicos para ejercer su función biológica.
Se eliminan por difusión y degradación.
Hasta hace poco se pensaba que la neurona producía 1 único tipo de NT. A día de hoy
hay muchos tipos de neuronas que sintetizan y liberan 2 o más neurotransmisores,
pudiendo coexistir en la misma neurona un NT de molécula pequeña con un NT de
molécula grande. A estos se les denomina cotransmisores.
COTRANSMISORES
El glutamato (molécula pequeña) puede coexistir con la sustancia P.
El GABA (pequeña) puede coexistir con la sustancia P.
La glicina (pequeña) con la neurotensina.
La Ach (pequeña) con la sustancia P, el péptido relacionado con el gen de la calcitonina
o el péptido intestinal vasoactivo.
La dopamina (pequeña) con la encefalina.
La noradrenalina (pequeña) con el péptido Y.
La adrenalina (pequeña) con la encefalina, el neuropéptido Y, y la sustancia P.
Serotonina (pequeña) con el neuropéptido Y, la encefalina o el péptido intestinal
vasoactivo.
RECEPTORES
La acción de un NT depende del tipo de receptor sobre el que actúa.
Receptores: proteínas de membrana importantes en la respuesta biológica que se
inician cuando se les une un NT.
2 grupos:
IONOTRÓPICOS O CANALES IÓNICOS ASOCIADOS A LIGANDO
Tienen un dominio extracelular, un dominio intracelular y varios dominios
transmembrana que se organizan para formar un poro, que forma el canal iónico para
permitir el flujo de iones del medio extracelular al intracelular y viceversa.
Este canal iónico tiene acoplado un receptor para la unión del NT. Cuando se une, el
canal se abre y permite el flujo de iones.
Si se abre un canal de sodio y potasio: entran sodio o potasio. Son cargas positivas,
por lo que aumenta el potencial de membrana, produciéndose excitación. Si esto
ocurre a nivel presináptico, se genera una facilitación presináptica, y si ocurre a
nivel postsináptico se genera un potencial excitatorio postsináptico.
Si el canal iónico es permeable al potasio y al cloro: entra cloro y sale potasio. Se
produce una hiperpolarización de la membrana. Si esto ocurre a nivel presináptico,
se genera una inhibición presináptica, y si ocurre a nivel postsináptico se genera un
potencial inhibitorio postsináptico.
La activación de receptores nicotínicos y receptores de serotonina 5HT3, permiten la
entrada de sodio a la neurona, excitando la membrana.
Otros receptores que producen activación son los AMPA, NDMA y Kainato, por el
glutamato, que permite el flujo de diferentes iones (sodio, potasio y calcio).
La activación de GABA y glicina produce inhibición, permitiendo la entrada de
cloro, produciendo una hiperpolarización.
METABOTRÓPICOS
Tienen un dominio extracelular, un dominio intracelular y varios dominios
transmembrana.
Acoplados a la proteína G, que una vez activada desencadena una cascada intracelular
implicando a segundos mensajeros, que generan canales iónicos de membrana para
producir hiperpolarización o despolarización.
Si la proteína G es de tipo S (proteína Gs) estimula la adenilato ciclasa, lo que
incrementa las concentraciones de AMPc y activación de la PKA, relacionada con la
activación de canales de potasio. Sale potasio fuera y se produce hiperpolarización.
Si la proteína G es de tipo I (proteína Gi) inhibe la adenilato ciclasa, y
una disminución de AMPc, inhibiendo la salida de potasio, por lo que
aumenta el potencial de membrana.
Si la proteína G es de tipo Q (proteína Gq), activa la PLC (fosfolipasa C), que actúa
sobre el inositol difosfato (PIP2) para estimular la síntesis de DAG e IP3. Estos
segundos mensajeros participan en la despolarización de la membrana.
Las respuestas metabotrópicas suelen ser más lentas que las de receptores
ionotrópicos, ya que los metabotrópicos implican una cascada intracelular con la
participación de 2os mensajeros, lo que ralentiza el proceso.
AUTORRECEPTORES
Receptor presináptico que inhibe la secreción adicional del NT, y proporciona un
control de retroalimentación.
EJEMPLO: tenemos una neurona adrenérgica. La liberación de
noradrenalina activa receptores postsinápticos, desencadenando
eventos intracelulares que aumentan el AMPc. Hay un receptor alfa2
adrenérgico en la membrana presináptica, que, al ser activado por la
noradrenalina, inhibe la liberación de más noradrenalina.
HETERORRECEPTORES
Receptor presináptico activado por un ligando diferente a lo que la
neurona sintetiza y secreta.
EJEMPLO: vemos una neurona colinérgica que libera Ach, y que activa
receptores muscarínicos M2 (autorreceptores), que inactivan la
adenilato ciclasa y por tanto a una disminución de AMPc. La liberación
de Ach es inhibida por el acoplamiento de noradrenalina liberada de
neuronas adrenérgicas.
En la membrana colinérgica se expresa un receptor adrenérgico alfa2, que es activado
por la noradrenalina de la neurona adrenérgica. Esa activación conlleva a una
inhibición de la liberación de Ach.
ACETILCOLINA
La Ach activa receptores muscarínicos o nicotínicos. Los nicotínicos son
146ionotrópicos, y los muscarínicos son receptores metabotrópicos a
proteínas G y pueden tener un efecto excitador o inhibidor dependiendo
del subtipo.
Si la Ach activa receptores M1, M3 y M5, estimulan la PLC, aumentando
las concentraciones intracelulares de IP3 y DAG y un incremento de la
entrada de calcio. Tienen un efecto excitador. Proteína Gq.
Si la Ach se une a receptores M2 y M4, disminuyen las concentraciones intracelulares
de AMPc, incrementa la salida de potasio y disminuye la entrada de calcio. Tiene un
efecto inhibidor. Los M2 y M4 se pueden localizar en las membranas presinápticas,
funcionando como autorreceptores o heterorreceptores. Cuando son activados por la
Ach, disminuyen la liberación del neurotransmisor.
Si la Ach se une a receptores nicotínicos, aumenta la entrada de sodio y calcio. Tiene
una función excitadora, y aumenta la liberación del NT, y tiene un papel en la
dependencia de la nicotina.
AMINOÁCIDOS
El GABA se puede unir a receptores ionotrópicos (GABAA). Si lo hace aumentan las
corrientes de entrada de cloro, por lo que tiene función inhibidora, disminuye el
potencial de membrana.
Si el GABA se une a receptores metabotrópicos (GABAB) disminuyen las
concentraciones de AMPc, aumenta la salida de potasio y disminuye la entrada de
calcio, por lo que tiene una función inhibidora.
La glicina se une a receptores sensibles a estricnina, aumentando la entrada de cloro,
por lo que tiene una función inhibidora, disminuye el potencial de membrana
La unión de glicina a receptores insensibles a estricnina tiene un efecto coagonista en
el receptor NMDA, por lo que una función excitadora.
El glutamato se puede unir a receptores ionotrópicos (NMDA, AMPA, KA), y provoca
un incremento de la entrada de sodio y calcio, por lo que tiene función excitadora.
Si el glutamato se une a receptores metabotrópicos (mglu1, mglu5), estimula la PLC,
por lo que aumenta el IP3, DAG y calcio, por lo que tiene función excitadora.
La unión del glutamato a receptores metabotrópicos (mglu2, mglu4, mglu6 y mglu8),
disminuye los niveles de AMPc, un aumento de la salida de potasio y una disminución
de la entrada de calcio. Tiene función inhibidora, produce hiperpolarización.
AMINAS BIÓGENAS
La dopamina se une a receptores metabotrópicos (d1 y d5), que estimulan la adenilato
ciclasa, por lo que aumentan los niveles de AMPc y PKA. Tiene función excitadora.
Si la dopamina se une a receptores metabotrópicos (d2, d3 y d4) disminuyen los
niveles de AMPc, aumenta la salida de potasio y disminuye la entrada de calcio. Tiene
función inhibidora.
La histamina se puede unir a receptores metabotrópicos (H1, H2 y H3).
- H1 estimula la PLC, por lo que aumentan los niveles de IP3, DAG y la entrada de
calcio. Tiene función excitadora.
- H2 estimula la AC, aumentando los niveles de AMPc y la PKA. Tiene función
excitadora.
- H3 funciona como autorreceptor y heterorreceptor. Disminuyen los niveles de
AMPc, aumenta la salida de potasio y disminuye la entrada de calcio, por lo que
tiene un efecto inhibidor.
NORADRENALINA
Se puede unir a receptores alfa adrenérgicos o beta adrenérgicos.
Los receptores metabotrópicos alfa1 estimulan la PLC, por lo que aumentan los
niveles de IP3, DAG y la entrada de calcio. Tiene función excitadora.
Los receptores alfa2 disminuyen los niveles de AMPc. Tiene efecto inhibidor.
Los receptores beta1 y beta2 están acoplados a proteínas Gs, por lo que se estimula la
AC, y aumentan los niveles de AMPc y PKA. Tienen función excitadora sobre el cortex
del sistema límbico y del núcleo accumbens.
SEROTONINA
Se puede unir tanto a receptores metabotrópicos (5-HT1, 5-HT2 y 5-HT4) como
ionotrópicos (5-HT3).
Si se une a receptores 5-HT1 disminuyen los niveles de AMPc. Tiene función
inhibidora.
Si se une a receptores 5-HT2, acoplado a la proteína Gq, estimula la PLC, aumentando
el IP3, DAG y la entrada de calcio. Tiene función excitadora.
Si se une a receptores 5-HT3 aumentan los niveles de sodio y calcio. Tiene función
excitadora.
Si se une a receptores 5-HT4 aumentan los niveles de AMPc y PKA, por lo que tiene
función [Link]
NEUROPÉPTIDOS
Los péptidos opioides disminuyen los niveles de AMPc, aumenta la salida de potasio y
disminuye la entrada de calcio. Tiene función inhibidora.
Las taquicininas activan receptores metabotrópicos (NK1, NK2, NK3) acoplados a
proteínas Gq, por lo que estimulan la PLC, aumentando los niveles de IP3, DAG y la
entrada de calcio. Tiene función excitadora.
DOPAMINA
NT del grupo de las catecolaminas. Es el NT catecolaminérgico más importante del
SNC.
Controla una gran variedad de funciones: modulación de la actividad sensorial, control
de la actividad motora y actividad endocrina, y participa en los procesos de
aprendizaje, memoria, emotividad, afectividad y motivación.
Alteraciones en el sistema dopaminérgico derivan en múltiples enfermedades
neurológicas: Parkinson y enfermedad de Huntington.
Relacionado con el desorden de déficit de atención (hiperactividad) y con varias
patologías endocrinas.
Las neuronas responsables de sintetizar dopamina se encuentran en
diferentes áreas del SNC:
- Sustancia negra.
- Área tegmental ventral (VTA).
- Hipotálamo.
- Cuerpo estriado.
- Varias partes de la corteza frontal.
VÍAS DOPAMINÉRGICAS
La distribución de neuronas dopaminérgicas da lugar a los sistemas dopaminérgicos:
- Sistema mesoestriatal.
- Sistema mesocorticolímbico.
- Sistema tuberoinfundibular.
Las vías dopaminérgicas no son más que extensiones de los axones de las neuronas
dopaminérgicas:
1. Vía nigroestriatal: el soma de las neuronas se encuentra en la sustancia negra
compacta del mesencéfalo. Sus axones se proyectan a la parte dorsal del cuerpo
estriado. Esta vía está asociada al control motor, por lo que alteraciones en ella están
relacionadas con la enfermedad de Parkinson y disquinesias de la enfermedad de
Huntington.
2. Vía mesolímbica: el soma se encuentra en el área tegmental ventral del
mesencéfalo. Sus axones se extienden a la amígdala y al núcleo
de accumbens. Relacionada con respuestas motivacionales y
afectivas (esquizofrenia, depresión).
3. Vía mesocortical: formada por neuronas dopaminérgicas que
se extienden en el área tegmental ventral hacia la corteza
central. Importante en la función cognitiva, y está relacionada
respuestas relacionadas con la motivación. Está relacionada con
la vía mesolímbica.
4. Vía tuberoinfundibular: proyecciones de neuronas desde el núcleo arcuato del
hipotálamo basal, y se extiende hasta la región infundibular. Participa en la regulación
de la secreción de prolactina en la adenohipófisis, inhibiendo su liberación.
SÍNTESIS Y LIBERACIÓN DE DOPAMINA
Tiene lugar en las terminaciones nerviosas dopaminérgicas, y sigue una cascada
común a las demás catecolaminas.
Tiene como precursor la tirosina (altas concentraciones en sangre), la cual es
recaptada por la terminación nerviosa. En el citosol de la terminación nerviosa sufre
una hidroxilación y una descarboxilación.
- La hidroxilación está mediada por la tirosina hidroxilasa (TH) y requiere como
factor tetrahidrobiopterina (BH4). Se forma la L-dopa.
- L-dopa es descarboxilada por la L-dopa descarboxilasa a dopamina.
La dopamina es empaquetada en vesículas. La liberación de la dopamina es
dependiente de calcio. La llegada de un PA abre canales de calcio dependientes de
voltaje, permitiendo el flujo de calcio desde el exterior al interior de la célula.
Se forma el complejo calcio-calmodulina, esencial para que la vesícula se fusione con
la terminación nerviosa. Una vez fusionada se libera la dopamina a la hendidura
sináptica, y activa los receptores postsinápticos para ejercer su efecto biológico.
Parte de la dopamina es recaptada por la neurona presináptica por transportadores.
Esta dopamina recaptada inhibe la tirosina hidroxilasa (retroalimentación negativa).
La dopamina puede activar autorreceptores: receptores
presinápticos dopaminérgicos que inhiben la liberación
de más dopamina.
Hay heterorreceptores como el GABAB, que inhibe la
liberación de dopamina cuando es activado por el GABA.
Hay heterorreceptores NMDA (glutamato) y adenosina
(A2), que estimulan la liberación de dopamina.
La dopamina recaptada puede ser convertida en ácido
dihidroxifenilacético (DOPAC), el cual difunde a través de la membrana en la
hendidura sináptica, y por la enzima catecol-o-metiltransferasa (COMT) se convierte
en ácido homovanílico (HVA).
RECEPTORES DOPAMINÉRGICOS
Se han descrito 5 tipos de receptores de dopamina, que han sido agrupados en 2
familias principales: D1 y D2. Los 5 receptores son metabotrópicos, por lo que están
acoplados a proteínas G.
a) D1: formado por el receptor D1 y D5. Están acoplados a la proteína Gs, y cuando
son activados por la dopamina estimulan la adenilato ciclasa, que aumenta los niveles
intracelulares de AMPc y la PKA. La PKA fosforila la proteína fosfatasa neuronal
regulada por dopamina (DARPP-32) y AMPc, e inhibe la fosfatasa 1 (PP-1). La PKA, al
actuar sobre canales iónicos de sodio y calcio, produciendo la entrada de calcio tipo L y
la excitación de la membrana. D5 tiene más afinidad para la dopamina que D1.
- D1 es el receptor dopaminérgico más abundante del SNC, y se
encuentra en varias estructuras del encéfalo. Tiene un papel
importante en la locomoción, aprendizaje, memoria, atención, control
del impulso, sueño y regulación renal.
- D5 se ha identificado en el hipocampo, en el tálamo, en el cuerpo
estriado, en el hipotálamo y en la corteza frontal y temporal. Participa
en la cognición, atención, toma de decisiones, aprendizaje motor y secreción de
renina.
b) D2: formada por receptores D2, D3 y D4. Están acoplados a la proteína Gi. Cuando
se activan inhiben la adenilato ciclasa, por lo que disminuye el AMPc y la PKA.
Producen inhibición.
- D2: se expresan en el cuerpo estriado, en el área tegmental ventral
del mesencéfalo y en la corteza cerebral. Implicado en locomoción,
aprendizaje, memoria, atención, regular el sueño y comportamiento
reproductivo.
- D3: en el cuerpo estriado y en la corteza cerebral. Implicado en
locomoción, cognición, atención, control del impulso y regulación del
sueño e ingesta de comida.
- D4: en la corteza frontal, la amígdala, hipotálamo y núcleo accumbens.
Participa en cognición, control de impulsos, atención, regulación del sueño y el
comportamiento reproductivo.
ACETILCOLINA
Primer NT identificado.
Principal NT de las neuronas bulboespinales y se encuentra en los
núcleos basales, en algunas zonas de la corteza motora, en el
tálamo, en el hipotálamo, en el hipocampo y en las áreas límbicas.
Es un NT importante también en el sistema nervioso periférico
(SNP): está presente en las fibras somáticas, en todas las neuronas
preganglionares del SNA y en las neuronas preganglionares del
sistema nervioso parasimpático.
Las vías colinérgicas con axones largos no siempre tienen sus cuerpos celulares en
núcleos bien definidos. Se observan proyecciones que salen del complejo
pontomesencefalotegmentario, que se extienden hasta el tálamo, el hipotálamo y los
núcleos septales y basales (núcleo basal de Meynert). De ahí salen proyecciones que
van al hipocampo, a la corteza y al bulbo olfatorio.
La Ach promueve la interconexión entre neuronas, fomentando la consolidación de
recuerdos, del aprendizaje, la memoria, la motivación y la capacidad de atención.
Alteraciones en la síntesis y liberación del metabolismo colinérgico o algún daño en las
vías colinérgicas genera un déficit en estas funciones.
El núcleo basal de Meynert es una de las zonas que más daños sufre: por el Alzheimer,
Parkinson y demencia por cuerpos de Lewy.
Participa en la regulación de los ciclos del sueño. La serotonina estimula la fase REM.
SÍNTESIS Y LIBERACIÓN DE ACETILCOLINA
La Ach se sintetiza a partir de 2 precursores:
- Colina: procede de la colina circulante, producida en hígado, de la
fosfatidilcolina y de la colina que resulta de la degradación de la Ach cuando es
liberada. La colina es recaptada por un transportador de colina para formar
parte de la nueva síntesis de Ach.
- Acetil-CoA: del metabolismo de la mitocondria de la neurona colinérgica.
Está mediada por la colinoacetil transferasa (ChAT).
Una vez se sintetiza la Ach, es almacenada en las vesículas, donde se mantiene estable,
porque la Ach es muy lábil, y se convierte fácilmente en colina.
La liberación de Ach depende del calcio. La llegada de un PA activa
canales de calcio neuronales que producen la entrada de calcio del
espacio extracelular al citosol. Este calcio se une a la calmodulina
para desplazar la vesícula a la membrana del botón terminal. La
vesícula se fusiona para liberar la Ach.
Parte de esta Ach activa receptores colinérgicos sinápticos para
ejercer su efecto biológico.
Otra parte se degrada por la acetilcolinesterasa (AChE) para formar acetato y colina.
Esta colina es recaptada a través de un transportador de colina.
Cuando la Ach se libera activa receptores presinápticos:
- M2: funciona como autorreceptor. Una vez activado, inhibe la liberación de
más Ach.
- En la membrana presináptica de la neurona colinérgica existen
heterorreceptores como los receptores adrenérgicos alfa2, que cuando son
activados por noradrenalina, inhiben la liberación de más Ach.
- Receptores nicotínicos ionotrópicos: facilitan la liberación de más Ach.
RECEPTORES COLINÉRGICOS
Se dividen en 2 grandes grupos:
a) Nicotínicos: son receptores ionotrópicos. Permiten el flujo de iones
de sodio y calcio. Producen una excitación de la membrana.
b) Muscarínicos: son receptores metabotrópicos. Están acoplados a proteínas G
- M2 y M4: acoplados a la proteína Gi, y cuando son activados por la Ach,
producen la inhibición de la adenilato ciclasa, disminuyendo el AMPc y por
tanto una disminución de la PKA. Producen inhibición, disminuyendo la
excitabilidad de la membrana.
- M1, M3 y M5: acoplados a la proteína Gq, y cuando son activados estimulan la
PLC, produciendo la hidrólisis de PIP2 a DAG e IP3. El IP3 activa receptores de
IP3 que se expresan en la membrana del retículo sarcoplásmico para producir
la liberación de calcio. Se produce excitación de la membrana.
NORADRENALINA
También conocida como norepinefrina. Es un NT de la familia de las catecolaminas, y
se encuentra extendida por todo el organismo, incluido el cerebro. Actúa como NT,
como neuromodulador y como hormona.
Desempeña un papel importante en el SNA, ya que está presente en las fibras
postganglionares simpáticas, regulando varios procesos fisiológicos periféricos.
A nivel del SNC, se le considera un NT de las neuronas del locus coerulus, un núcleo de
neuronas localizado en la protuberancia del tallo cerebral que envía eferencias a
regiones corticales y subcorticales como la médula espinal, el núcleo del raf medial,
núcleo de raf dorsal, formación reticular, hipocampo, corteza prefrontal, área motora
primaria, área somatosensorial, área olfatoria, núcleo subtalámico, sustancia negra y
cerebelo.
La inervación adrenérgica del límbico, sobre todo la amígdala, está involucrada en
estados de miedo, ansiedad y fobias.
Las proyecciones adrenérgicas que van a la corteza frontal están
relacionadas con el estado de ánimo y la atención.
La proyección adrenérgica que va al hipotálamo se une a las proyecciones
dopaminérgicas y forman el haz prosencefálico medial, relacionado con
procesos de recompensa.
La inervación del hipotálamo controla la toma de alimento por la inervación del centro
de la saciedad, y participa en la regulación neuroendocrina del hipotálamo.
La inervación adrenérgica del cerebelo se relaciona con el control de los movimientos
motores, en especial con los temblores.
La noradrenalina inhibe el sueño, regulando el sueño-vigilia. Hace sinapsis inhibitorias
con neuronas colinérgicas inductoras del sueño, responsables del paso al sueño REM.
La noradrenalina bloquea el paso al sueño REM
La noradrenalina también está controlada por las neuronas gabaérgicas.
NORADRENALINA-ADRENALINA
Es el metil derivado de la noradrenalina, y se secreta en la médula suprarrenal.
La síntesis de la adrenalina tiene como precursor la tirosina, y sigue la vía de síntesis
de la dopamina y noradrenalina. Después de la síntesis de noradrenalina, la enzima
fentolamina N-metiltransferasa (FNMT) convierte la noradrenalina en adrenalina.
La adrenalina es un mediador en las terminaciones nerviosas simpáticas
postganglionares.
La noradrenalina y adrenalina cuando son sintetizadas en la célula
cromafín, son liberadas directamente al torrente sanguíneo, y de ahí
recorren un camino hasta alcanzar el órgano diana para ejercer su efecto.
La adrenalina realiza sus funciones sobre diversos órganos y sistemas,
facilitando la transmisión del impulso nervioso de una fibra a otra. Actúa también
sobre músculo, tejido adiposo, sobre hígado, y prepara al organismo para la lucha,
poniéndole en guardia para una situación de estrés.
Entre sus efectos fisiológicos destaca el aumento de concentración de glucosa en
sangre, el aumento de la tensión arterial, el aumento del ritmo cardiaco, la dilatación
de las pupilas y la dilatación de los conductos de aire.
SÍNTESIS Y LIBERACIÓN DE NORADRENALINA
La noradrenalina es una catecolamina, por lo que su síntesis es común a la de la
dopamina.
Tiene como precursor la tirosina (altas concentraciones en sangre), la cual es
recaptada por la terminación nerviosa. En el citosol de la terminación nerviosa sufre
una hidroxilación y una descarboxilación.
- La hidroxilación está mediada por la tirosina hidroxilasa (TH) y requiere como
factor tetrahidrobiopterina (BH4). Se forma la L-dopa.
- L-dopa es descarboxilada por la L-dopa descarboxilasa a dopamina.
La dopamina es captada por los granos de la terminación nerviosa debido a los
transportadores vesiculares de monoaminas, que internalizan la dopamina en las
vesículas, y es donde se encuentra la dopa beta-hidroxilasa, que
convierte la dopamina en noradrenalina.
La noradrenalina es almacenada en vesículas hasta que llega un PA
que produce la entrada de calcio, movilizándose la vesícula hasta
que se fusiona con la membrana de la terminación nerviosa
adrenérgica.
La noradrenalina se libera por exocitosis, y activa receptores
adrenérgicos postsinápticos, ejerciendo su efecto biológico.
Parte de la noradrenalina va a ser degradada por la catecol o-metiltransferasa
(COMT), que se encuentra en la membrana de los tejidos postsinápticos, y forma el
dihidroxifeniletilenglicol (DOPEG).
Otra parte de la noradrenalina es recaptada a la neurona presináptica a través de
transportadores de catecolaminas. Una vez en el citosol del axón, la noradrenalina
puede ser receptada por vesículas presinápticas, utilizando transportadores con gasto
de energía, o pueden ser oxidadas por la acción de monoaminaooxidasas.
La liberación de noradrenalina se ve afectada por la activación de receptores
presinápticos, ya sean:
- Autorreceptores como alfa2, que inhibe más noradrenalina.
- Heterorreceptores como M2, que cuando son activados por la Ach de las
neuronas colinérgicas inhiben la liberación de más noradrenalina.
- Otros heterorreceptores son los dopaminérgicos de la familia D2, que
disminuyen la liberación de noradrenalina.
- Hay heterorreceptores opioides, que al ser siempre inhibidores, su respuesta
siempre va a implicar un descenso de liberación de noradrenalina.
RECEPTORES ADRENÉRGICOS
Son metabotrópicos, por lo que están asociados a proteínas G. Receptores alfa
adrenérgicos (alfa1 y alfa2) y receptores beta adrenérgicos (beta1 y beta2).
Alfa1: acoplados a una proteína Gq. Estimulan la PLC, que forma IP3 y DAG como
segundos mensajeros. IP3 produce una movilización de calcio, y DAG activa la PLC, que
produce fosforilación de canales iónicos y proteínas de la membrana. Excitación.
Alfa2: acoplados a proteínas Gi, por lo que son inhibidores, disminuyen la actividad de
la adenilato ciclasa, bajando las concentraciones de AMPc y disminuyendo la actividad
de la PKA, por lo que baja la permeabilidad de canales de calcio, y facilita la apertura
de canales presinápticos de potasio.
Los receptores beta adrenérgicos (beta1 y beta2) son receptores metabotrópicos
asociados a proteínas Gs, por lo que estimulan la actividad de la adenilato ciclasa,
aumentando los niveles de AMPc, con la activación de la PKA. Esto produce excitación.
Los receptores alfa adrenérgicos:
- Alfa1: más afinidad por la adrenalina que noradrenalina.
- Alfa2: más afinidad por la noradrenalina que por la adrenalina.
Los receptores beta adrenérgicos:
- Beta1: misma afinidad tanto para la adrenalina como noradrenalina.
- Beta2: más afinidad por la adrenalina que por la noradrenalina.
- Beta3: más afinidad por la noradrenalina que por la adrenalina.
SEROTONINA
NT y neuromodulador implicado en la regulación de estados de ánimo y funciones
fisiológicas y conductas. Regula la conducta social, la conducta sexual, la atención, la
ansiedad, los patrones rítmicos (locomoción, masticación y respiración) y el sueño.
Alteraciones en el sistema serotoninérgico están relacionadas con trastornos
conductuales y neurológicos como la depresión, trastornos alimenticios,
esquizofrenia, epilepsia y ansiedad.
Los somas de las neuronas serotoninérgicas se encuentran agrupados en los
núcleos de raf en el tallo cerebral, e inervan todo el SNC.
Los axones de los núcleos más posteriores ascienden para inervar el encéfalo,
incluyendo los ganglios basales, el tálamo, la corteza y el cerebelo.
Los axones de los núcleos anteriores descienden para inervar la médula espinal.
SÍNTESIS Y LIBERACIÓN DE SEROTONINA
Se sintetiza en la terminación nerviosa serotoninérgica y tiene como precursor el
triptófano, que es captado a la terminación nerviosa por un transportador de L-
aminoácidos (aa) aromático dependiente de sodio.
Una vez ahí, ocurre la hidroxilación del triptófano a 5-hidroxitriptófano por acción de
la triptófano hidroxilasa.
El 5-hidroxitriptófano es descarboxilado por la enzima descarboxilasa
L-aa aromático para formar la serotonina (5-hidroxitriptamina).
La serotonina es empaquetada en las microvesículas sinápticas por un
transportador, y queda almacenada hasta que entra calcio. La llegada
de un PA produce la apertura de canales de calcio, esencial para
movilizar la vesícula, y hacer que se fusione con la membrana de la
neurona presináptica.
Una vez ocurre la fusión, la serotonina es liberada por exocitosis en la hendidura
sináptica, y activa receptores serotoninérgicos postsinápticos para ejercer su efecto
biológico.
La serotonina también activa receptores presinápticos y autorreceptores
serotoninérgicos para inhibir más serotonina.
Parte de la serotonina puede difundir fuera de la hendidura sináptica, pero una parte
es recaptada por los transportadores de serotonina de la membrana presináptica.
La serotonina recaptada puede ser secuestrada por la microvesículas sinápticas, y
también puede ser inactivada por la monoaminaooxidasa para formar el 5-
hidroxiindol acetaldehído, que es el principal metabolito de la serotonina, que se
libera en orina e indica la tasa de serotonina en el organismo.
RECEPTORES SEROTONINÉRGICOS
Todos son receptores metabotrópicos, por lo que están asociados a proteínas G,
excepto el receptor 5-HT3, que es un receptor ionotrópico.
5-HT1a es un receptor metabotrópico asociado a la proteína Gi, por lo que disminuye
los niveles de AMPc. Tiene un efecto presináptico a nivel tanto somatodendrítico en
los núcleos del rad y a nivel terminal en el sistema límbico. Tiene un efecto inhibitorio.
5-HT2 está acoplado a la proteína Gq, por lo que activa la PLC e incrementa el IP3 y
DAG. Tiene efecto excitador.
- 5-HT2a se encuentra localizado en la corteza y en el estriado.
- 5-HT2c se encuentra en los plexos coroideos, donde regula el volumen y la
composición de líquido cefalorraquídeo.
5-HT3 son receptores ionotrópicos facilitadores y permiten la entrada de sodio y
potasio. Se localiza en los núcleos del tronco encefálico y en la medula espinal.
Funcionan como heterorreceptores en las neuronas gabaérgicas, dopaminérgicas y
colinérgicas, donde facilitan la liberación de NT. En el área próxima del tronco
encefálico, su liberación libera dopamina, mientras que en la médula espinal media
efectos nociceptivos (dolor), liberando la sustancia P.
5-HT4, 5-HT6 y 5-HT7 están acoplados a proteínas Gs. Aumentan las concentraciones
de AMPc, por lo que tienen un efecto excitador.
5-HT4 se encuentra como heterorreceptor en neuronas dopaminérgicas e hipocampo,
facilitando procesos cognitivos como la memoria a corto plazo y a largo plazo.
5-HT6 tiene importancia en el comportamiento emocional, ya que está presente en el
núcleo accumbens, los núcleos olfatorios, el estriado y la corteza. Casi todos los
fármacos psicotrópicos tienen a 5-HT6 como diana, y tiene afinidad por el SB.
5-HT7 se localiza en el núcleo supraquiasmático hipotalámico, donde está implicado
en la regulación de ritmos circadianos.
HISTAMINA
Se sintetiza en las neuronas histaminérgicas. Los somas se encuentran en los núcleos
tuberomamilares del hipotálamo posterior, y de ahí se proyectan por vías
descendentes y ascendentes.
Las vías ascendentes inervan todos los núcleos del hipotálamo, el septum lateral, el
estriado, el córtex y el sistema límbico, con gran inervación del hipocampo y del núcleo
accumbens. También hay proyecciones a la sustancia negra.
Las vías descendentes representan las proyecciones que inervan el bulbo y la médula.
El sistema histaminérgico está implicado en funciones como el control del
apetito, ya que la histamina suprime el apetito al activar el centro de la
saciedad a través de receptores H1.
El hipotálamo regula la secreción de la hormona adenocorticotropa (ACTH)
y de beta endorfinas que participan en la sensación de placer, y participa
en la regulación de hormonas hipofisarias como la oxitocina y la vasopresina.
También está implicado en el ciclo sueño-vigilia. Esto explica el efecto sedante de los
antihistamínicos, que bloquean los receptores H1, y la histamina no mantiene el
estado de vigilia.
Interviene también en la función antinociceptiva producida a través de receptores H1
y H2. La histamina media el efecto de la morfina (opioides). Los opioides liberan
histamina, que se une a receptores histaminérgicos H2, y disminuye la percepción
dolorosa.
SÍNTESIS Y LIBERACIÓN DE HISTAMINA
A nivel central se produce en las neuronas histaminérgicas en un solo paso de
descarboxilación a partir de la L-histidina, que accede al botón por transportador de L-
aminoácidos (aa).
Ahí, la L-histidina se descarboxila por acción de la histidina descarboxilasa, y se forma
la histamina. A medida que se sintetiza, la histamina es introducida en vesículas para
evitar su degradación. El empaquetamiento se da mediante transportadores
vesiculares de monoaminas (como en el resto). El transportador realiza un simporte
con hidrógeno, que implica la acción de una ATPasa.
La liberación de histamina se da por exocitosis a través de mecanismos
dependientes de calcio (como el resto). Una vez liberada en la hendidura
sináptica la histamina activa los receptores histaminérgicos
postsinápticos y puede activar autorreceptores como H3, que inhibe la
liberación de más histamina.
La histamina es eliminada por la diamino oxidasa (DAO), que convierte
la histamina en ácido acético.
La histamina también puede ser convertida en tele-metilhistamina por la histamina-N-
metil-transferasa (HMT). Esta enzima degrada histamina en el tejido hepático, pero
también se expresa en otros tejidos.
H3 son heterorreceptores que se expresan en neuronas adrenérgicas,
serotoninérgicas, dopaminérgicas, gabaérgicas y glutamatérgicas. Inhiben la liberación
de sus NT.
RECEPTORES HITAMINÉRGICOS
Todos son metabotrópicos.
El H1 está asociado a una proteína Gq, por lo que estimula la PLC, aumentando IP3 y
DAG. La activación de estos segundos mensajeros excita la membrana.
H3 y H4 están asociados a proteínas Gi, que inhiben la adenilato ciclasa, disminuyendo
las concentraciones de AMPc y la PKA, produciendo inhibición (hiperpolarización).
H2 está asociado a una proteína Gs, por lo que activa la adenilato ciclasa, aumentando
los niveles de AMPc y PKA, produciendo excitación.
HASTA AQUÍ LAS AMINAS BIÓGENAS.
NEUROTRANSMISORES AMINOÁCIDOS
Se dividen en 2 grupos:
- NT inhibitorios: GABA y glicina.
- NT excitatorios: glutamato.
GLUTAMATO
Es un aa distribuido por todo el SNC, donde funciona como NT excitador. Es el
principal NT excitador del SNC, interviene en el 75% de la transmisión de impulsos
excitadores.
Algunas de las vías principales son:
- Vía cortico-talámica.
- Vía tálamo-cortical.
- Vía hipocampal-estriatal.
- Proyección cortico-troncoencefálica.
- Vía cortico-estriatal.
- Vía cortico-corticales.
Participa en funciones como el control motor, el control de las emociones, procesos
cognitivos de aprendizaje y memoria, plasticidad durante el desarrollo embrionario y la
vida adulta.
SÍNTESIS Y LIBERACIÓN DE GLUTAMATO
Se puede dar por 2 vías:
1. A través de la transaminación del alfa-cetoglutarato. El alfa-cetoglutarato
producido en el ciclo de Krebs se convierte en glutamato por acción de la enzima
GABA-aminotransferasa.
2. Por la vía glutamina: la glutamina se hidroliza a glutamato por la glutaminasa.
El glutamato es captado por las vesículas presinápticas por un transportador de
glutamato vesicular. Una vez sintetizado y almacenado, se libera por exocitosis en la
hendidura sináptica y por mecanismos dependientes de calcio, y activa receptores
glutamatérgicos postsinápticos para ejercer su efecto biológico.
Parte del glutamato es recaptado por la neurona presináptica
glutamatérgica.
Parte del glutamato se puede reducir a glutamina por acción
de la glutamina sintetasa en la astroglía.
El glutamato de la hendidura sináptica puede ser recaptado
por astrocitos (astroglía) a través de transportadores que
introducen el glutamato dentro de la célula, y por la
glutamina sintetasa se transforma glutamina, la cual viaja
hacia la neurona presináptica glutamatérgica, donde participa en la síntesis de nuevo
glutamato.
RECEPTORES GLUTAMATÉRGICOS
RECEPTORES IONOTRÓPICOS
Median respuestas excitadoras. Se denominan en función de su agonista sintético:
- Receptores kainato: afinidad para el ácido kaínico (empleado para
epilepsia).
- Receptor AMPA: afinidad para el aminohidroximetilisosazolpropiónico.
- Receptor NMDA: afinidad para el N-metil D-aspartato.
La activación de los receptores AMPA genera el componente precoz del PA. Se les
denomina canales de baja conductancia, y son permeables al sodio y al potasio.
La activación de los receptores NMDA genera un PA postsináptico lento, y abre
canales de alta conductancia permeables al calcio.
RECEPTORES METABOTRÓPICOS
Median respuestas tanto activadoras como inhibidoras.
- Grupo 1: receptores de membrana postsinápticos. Constituido por el receptor
glutamatérgico 1 y el receptor glutamatérgico 5.
- Grupo 2: receptores presinápticos. Constituido por los receptores 2 y 3.
- Grupo 3: receptores presinápticos. Constituido por los receptores 4, 6 y 7.
Estos receptores del grupo 1 están acoplados a proteínas Gs que estimulan el AMPc,
pero también Gq que estimulan el IP3 y DAG, produciendo activación.
Los receptores del grupo 2 están acoplados a proteínas Gi que disminuyen el AMPc,
produciendo inhibición.
Los receptores del grupo 3 están asociados a proteínas Gi que disminuyen también el
AMPc, produciendo inhibición.
GABA
Es el ácido gamma-aminobutírico. Es el principal NT del SNC. Se encuentra en muy
grandes concentraciones y de forma difusa en varias áreas.
Las sinapsis gabaérgicas son conexiones de interneuronas o neuronas de axones
cortos. Hay 4 vías:
- Del estriado a la sustancia negra.
- De la sustancia negra al tálamo.
- Del núcleo tegmental ventral a la corteza prefrontal.
- De las células de Purkinje al núcleo vestibular y al cerebelo.
El sistema límbico está inervado por el sistema gabaérgico. Se han
detectado proyecciones en el hipocampo, hipotálamo, bulbo olfatorio y corteza.
También está presente en el bulbo del tallo cerebral.
Tiene un papel importante en el control del comportamiento motor. Alteraciones en
este sistema provocan patologías motoras como el Parkinson, la discinesia tardía o la
corea de Huntington.
El sistema gabaérgico instaura el sueño REM (como la acetilcolina, no como la
noradrenalina y la histamina), manteniendo inactivas a las noradrenérgicas y
serotoninérgicas.
SÍNTESIS Y LIBERACIÓN DE GABA
Se sintetiza en las terminaciones nerviosas gabaérgicas a partir del glutamato, cuyo
precursor es la glucosa.
En las neuronas gabaérgicas la glutamato descarboxilasa convierte el glutamato en
GABA, que es empaquetado en vesículas sinápticas por un transportador especifico de
aa inhibidores.
Su liberación se da por exocitosis como en los otros. Una vez liberado en la hendidura
sináptica, activa receptores gabaérgicos para producir su efecto en las neuronas
postsinápticas.
Parte del GABA es recaptado a través de transportadores asociados al sodio de alta
afinidad localizados en la membrana presináptica y en los astrocitos.
En astrocitos el GABA es metabolizado gracias a la GABA transaminasa, generando el
succinil semialdehído.
La succinil semialdehído deshidrogenasa oxida el succinil semiladehído al succinato.
El succinato entra en el ciclo de Krebs, donde forma el alfa-cetoglutarato, y se obtiene
la síntesis de glutamina.
La glutamina abandona los astrocitos hacia la neurona
presináptica gabaérgica para participar en la síntesis de nuevas
moléculas de GABA.
El GABA es recaptado para ser conducido a las vesículas
presinápticas por transportadores vesiculares de GABA.
El GABA puede ser metabolizado a glutamina por el mismo
proceso que ocurría en el astrocito. La glutamina participa en la
formación de glutamato y después de más GABA.
RECEPTORES GABAÉRGICOS
RECEPTORES IONOTRÓPICOS
- GABAA
- GABAC
Son postsinápticos, y cuando son activados por GABA, permiten la entrada de cloro,
produciendo hiperpolarización. Los agonistas de estos receptores son útiles como
inductores del sueño y relajación.
Cuando las concentraciones de cloro son elevadas (durante el desarrollo), la
activación de los canales produce corrientes de salida de cloro y una despolarización.
RECEPTORES METABOTRÓPICOS
- GABAB1: contiene el dominio extracelular con el sitio de unión para
el GABA.
- GABAB2: contiene el acoplamiento a la proteína Gi, que inhibe la
adenilato ciclasa, produciendo inhibición.
El receptor GABAB es un heterodímero con 2 diferentes subunidades de 7
dominios transmembrana cada uno, que se encuentran unidos por sus terminales
carboxilo. Se encuentran ubicados tanto en la membrana presináptica como
postsináptica, y pueden funcionar como:
- Autorreceptores: su activación inhibe el canal de calcio dependiente de
voltaje, reduciendo la entrada de calcio, lo que disminuye la liberación del NT.
- Heterorreceptores: la activación de neuronas glutamatérgicas disminuye la
liberación de glutamato.
HASTA AQUÍ LOS AMINOACIDOS.
PURINAS
ATP Y ADENOSINA
NT sintetizados y liberados desde las neuronas colinérgicas y desde las células foliares.
ATP
El ATP puede coexistir en vesículas con las catecolaminas. Actúa como un NT excitador
en las neuronas motoras de la médula espinal de los ganglios sensitivos y
autonómicos. También tiene acciones postsinápticas sobre neuronas del asta dorsal y
del hipocampo.
La actividad colinérgica está implicada en los procesos de memoria y aprendizaje, así
como en gestión de locomoción y control de la alimentación y patrones de sueño.
El ATP neuronal se produce por fosforilación oxidativa de las mitocondrias, utilizando
como precursor glucosa extracelular, ya que las neuronas tienen una capacidad muy
limitada de almacenamiento de glucosa.
En ausencia de glucosa, sustancias como el piruvato y lactacto son utilizadas para la
respiración mitocondrial, aunque su disponibilidad es breve.
Las ATPasas dependientes de hidrógeno de la membrana de la vesícula presináptica
utilizan ATP producido en el terminal presináptico. Bombean un ion de hidrógeno
hacia el interior de la vesícula, produciendo una acidificación del lumen y creando un
gradiente de protones que hace que entre el ATP.
El ATP es utilizado para la exocitosis de la vesícula. Una vez liberado el ATP, activa
receptores ionotrópicos o metabotrópicos para ejercer su efecto biológico en la
membrana postsináptica.
ADENOSINA
La adenosina resulta de la degradación del ATP. La ATPasa
convierte el ATP en ADP, y el 5-nucleotidasa convierte el 5-AMP
en adenosina.
La adenosina activa receptores presinápticos y postsinápticos
para producir su efecto biológico.
La adenosina también puede ser producida en el interior de la
célula a partir de S-adenosilmetionina, que pasa a S-
adenosilhomocisteína, que por la S-adenosilhomocisteína hidrolasa, pasa a adenosina.
La adenosina intracelular NO es almacenada en vesículas, pero puede ser
transportada al espacio extracelular por un transportador de nucleótidos.
RECEPTORES PURINÉRGICOS
2 grandes familias:
1. Receptores P1: a los que se une la adenosina. Varios subtipos: A1, A2A, A2B y A3.
2. Receptores P2: a los que se unen el resto de nucleótidos. Varios subtipos:
- P2Y: junto con los P1, son receptores metabotrópicos.
- P2X: receptores ionotrópicos.
- P2Y12, P2Y13, P2Y14, A1 y A3 están acoplados a proteínas Gi, inhiben la
adenilato ciclasa y disminuyen el AMPc y la PKA.
- P2Y11, A2A y A2B están acoplados a proteínas Gs, por lo que estimulan la
adenilato ciclasa, el AMPc y PKA.
- P2Y1, P2Y2, P2Y4, P2Y6 y P2Y11 están acoplados a proteínas Gq, que
estimulan la PLC, incrementando la formación de IP3 y DAG.
HASTA AQUÍ LAS PURINAS.
NEUROPÉPTIDOS OPIOIDES
ENDORFINAS Y ENCEFALINAS
NT opioides producidos en el SNC, y regulan el dolor, tanto a nivel espinal como
supraespinal.
En las vías ascendentes encontramos el sistema opioide en la médula y el tálamo.
En las vías descendentes encontramos el opioide en la sustancia gris periacueductal y
en zonas límbicas como la amígdala, el córtex, el hipocampo y en el occus cerelose,
donde se encuentran la mayor parte de receptores opioides, sobre todo en las
neuronas noradrenérgicas que se proyectan al hipocampo y a la amígdala.
El sistema opioide se puede localizar también en áreas de control motor como el
estriado.
Las endorfinas son polipéptidos largos que se secretan de un único precursor:
proopiomelanocortina (POMC), que sufre varios cortes peptídicos para formar la alfa,
beta y gamma endorfina.
La encefalina es una hormona peptídica formada por la unión de varios aa.
Estos péptidos no se forman en el citosol de las terminales presinápticos, sino que son
sintetizados por los ribosomas del soma neuronal, y forman parte de los NT de
moléculas grandes.
RECEPTORES OPIOIDES
Acoplados a proteínas Gi que actúan sobre canales de potasio, aumentando su
apertura e hiperpolarizando la célula, y sobre canales de calcio, disminuyendo su
apertura.
RECEPTOR MU (µ)
Formado por 2 subunidades: µ1 (mu1) y µ2 (mu2). Sus agonistas son las endorfinas.
Están presentes en médula, tálamo, sustancia gris periacueductal, amígdala y núcleo
del tracto solitario.
En la médula, la activación produce analgesia espinal, disminuyendo la liberación del
glutamato, por lo que disminuye la percepción dolorosa.
En el tálamo la activación produce analgesia supraespinal, disminuyendo la
percepción dolorosa.
En la sustancia gris periacueductal la activación produce una disminución del disparo
de neuronas gabaérgicas que inhiben las vías noradrenérgicas y serotoninérgicas
descendientes, inhibiendo el dolor.
En la amígdala produce euforia y bienestar, actuando a través del sistema
dopaminérgico a nivel de los fenómenos de recompensa.
En el núcleo del tracto solitario produce depresión respiratoria.
RECEPTORES K
3 subtipos: K1, K2 y K3. Su ligando es la dinorfina. Se distribuyen en el córtex, tálamo,
amígdala, hipocampo, hipotálamo y sustancia negra.
Su activación produce analgesia, disforia y sedación.
RECEPTOR GAMMA
2 subtipos: gamma1 y gamma2. Se unen a encefalinas. Se distribuyen en el córtex,
amígdala, puente, asta dorsal y medula espinal.
Su activación produce analgesia térmica.
MECANISMOS DE ACCIÓN
Estos receptores son metabotrópicos, acoplados a proteínas Gi, y producen inhibición
de la vía AC/AMPc/PKA, lo que conduce a un efecto analgésico.
HASTA AQUÍ LOS OPIOIDES.
TAQUICININAS
SUSTANCIA P
Polipéptido con 11 aa residuales que se encuentran en diversos tejidos del organismo,
incluido el SNC.
Involucrado en la percepción del dolor. Actúa tanto como neuromodulador como NT.
Se localiza en el encéfalo, médula espinal y en las protecciones sensoriales de dolor.
Función mayoritariamente excitatoria.
Es el NT más importante del haz paleoespinotalámico, que es el haz que transmite el
dolor lento crónico.
La sustancia P se une al receptor neuroquinina (NK), un receptor metabotrópico
acoplado a una proteína G. Su unión:
- Activa receptores NMDA.
- Inhibe canales de calcio dependientes de voltaje tipo N.
- Inhibición de canales de calcio activados por potasio.
También podría gustarte
- Neurotransmisores y Neuropéptidos en FisiologíaAún no hay calificacionesNeurotransmisores y Neuropéptidos en Fisiología32 páginas
- Sinapsis: Transmisión Nerviosa y TiposAún no hay calificacionesSinapsis: Transmisión Nerviosa y Tipos36 páginas
- Neurotransmisión y Sinapsis en PsicologíaAún no hay calificacionesNeurotransmisión y Sinapsis en Psicología4 páginas
- Fisiología del Sistema Nervioso: Sinapsis y Neurotransmisores100% (1)Fisiología del Sistema Nervioso: Sinapsis y Neurotransmisores28 páginas
- Neurotransmisores y NeuropeptidosAún no hay calificacionesNeurotransmisores y Neuropeptidos17 páginas
- Tipos y Funciones de Sinapsis NeuronalAún no hay calificacionesTipos y Funciones de Sinapsis Neuronal2 páginas
- Sinapsis y Neurotransmisores en el SNCAún no hay calificacionesSinapsis y Neurotransmisores en el SNC35 páginas
- Síntesis y Función de NeurotransmisoresAún no hay calificacionesSíntesis y Función de Neurotransmisores40 páginas
- Neurotransmisión y Sinapsis: Guía CompletaAún no hay calificacionesNeurotransmisión y Sinapsis: Guía Completa18 páginas
- Neurotransmisores y Neuropéptidos EsencialesAún no hay calificacionesNeurotransmisores y Neuropéptidos Esenciales15 páginas
- Neurotransmisores y Circulación SanguíneaAún no hay calificacionesNeurotransmisores y Circulación Sanguínea14 páginas
- Fisiología del Sistema Nervioso: Sinapsis y NeurotransmisoresAún no hay calificacionesFisiología del Sistema Nervioso: Sinapsis y Neurotransmisores27 páginas
- Neurotransmisores y Su Función en la SinapsisAún no hay calificacionesNeurotransmisores y Su Función en la Sinapsis51 páginas
- Funciones y Tipos de Células GlialesAún no hay calificacionesFunciones y Tipos de Células Gliales5 páginas
- Sinapsis y Neurotransmisores en FisiologíaAún no hay calificacionesSinapsis y Neurotransmisores en Fisiología10 páginas
- Sinapsis Química y NeurotransmisoresAún no hay calificacionesSinapsis Química y Neurotransmisores29 páginas
- Potencial de Acción y Sinapsis NeuronalAún no hay calificacionesPotencial de Acción y Sinapsis Neuronal76 páginas
- Funciones y Clasificación de NeurotransmisoresAún no hay calificacionesFunciones y Clasificación de Neurotransmisores20 páginas
- Sinapsis y Neurotransmisores ExplicadosAún no hay calificacionesSinapsis y Neurotransmisores Explicados20 páginas
- Estructura y Función del Sistema NerviosoAún no hay calificacionesEstructura y Función del Sistema Nervioso6 páginas
- Neurotransmisores y su función cerebralAún no hay calificacionesNeurotransmisores y su función cerebral63 páginas
- Sinapsis y Placa Neuromuscular: Guía CompletaAún no hay calificacionesSinapsis y Placa Neuromuscular: Guía Completa82 páginas
- Seminario 1-Neurotransmisores y NeuropeptidosAún no hay calificacionesSeminario 1-Neurotransmisores y Neuropeptidos25 páginas
- Sinapsis: Neurotransmisores y ReceptoresAún no hay calificacionesSinapsis: Neurotransmisores y Receptores28 páginas
- Neurotransmisores y Sinapsis en FisiologíaAún no hay calificacionesNeurotransmisores y Sinapsis en Fisiología17 páginas
- Tipos de Sinapsis y NeurotransmisoresAún no hay calificacionesTipos de Sinapsis y Neurotransmisores10 páginas
- Fisiología del Sistema Nervioso: Neuronas y SinapsisAún no hay calificacionesFisiología del Sistema Nervioso: Neuronas y Sinapsis4 páginas
- Neurotransmisores: Funciones y MecanismosAún no hay calificacionesNeurotransmisores: Funciones y Mecanismos16 páginas
- Fisiología de la Sinapsis NeuronalAún no hay calificacionesFisiología de la Sinapsis Neuronal3 páginas
- Funciones de la Noradrenalina en NeurotransmisiónAún no hay calificacionesFunciones de la Noradrenalina en Neurotransmisión29 páginas
- Neurotransmisión: Funciones y MecanismosAún no hay calificacionesNeurotransmisión: Funciones y Mecanismos7 páginas
- Introducción a los NeurotransmisoresAún no hay calificacionesIntroducción a los Neurotransmisores9 páginas
- Función y Clasificación de Neurotransmisores100% (4)Función y Clasificación de Neurotransmisores20 páginas
- Tipos de Sinapsis y NeurotransmisoresAún no hay calificacionesTipos de Sinapsis y Neurotransmisores56 páginas
- Mecanismos y Tipos de NeurotransmisoresAún no hay calificacionesMecanismos y Tipos de Neurotransmisores16 páginas
- M4.3 - Cuadro Comparativo y Esquemas Del Sistema LímbicoAún no hay calificacionesM4.3 - Cuadro Comparativo y Esquemas Del Sistema Límbico2 páginas
- Psicología Fisiológica: Emoción y ConductaAún no hay calificacionesPsicología Fisiológica: Emoción y Conducta9 páginas
- Mecanismos Neurobiológicos de la AdicciónAún no hay calificacionesMecanismos Neurobiológicos de la Adicción19 páginas
- Adicciones: Enfoque Holístico y FamiliarAún no hay calificacionesAdicciones: Enfoque Holístico y Familiar36 páginas
- Análisis de Artículos Periodísticos en Quinto GradoAún no hay calificacionesAnálisis de Artículos Periodísticos en Quinto Grado3 páginas
- Aprendizaje Cerebral en la ConducciónAún no hay calificacionesAprendizaje Cerebral en la Conducción15 páginas
- Resumen Motivacion Dopamina RecompensaAún no hay calificacionesResumen Motivacion Dopamina Recompensa3 páginas
- Práctica de Recuperación y RetenciónAún no hay calificacionesPráctica de Recuperación y Retención10 páginas
- Estímulos de Placer y Refuerzo en el CerebroAún no hay calificacionesEstímulos de Placer y Refuerzo en el Cerebro107 páginas
- Metilfenidato y su impacto inmunológicoAún no hay calificacionesMetilfenidato y su impacto inmunológico29 páginas
- Volkow, Nora D. Morales, M. (2015) : El Cerebro en Las Drogas: de La Recompensa A La AdicciónAún no hay calificacionesVolkow, Nora D. Morales, M. (2015) : El Cerebro en Las Drogas: de La Recompensa A La Adicción29 páginas
- Correlatos Neuronales de La Evocacion Emocional A Traves de La Musica (2014) - TraduccionAún no hay calificacionesCorrelatos Neuronales de La Evocacion Emocional A Traves de La Musica (2014) - Traduccion20 páginas
- Montessori y Atención Focalizada InfantilAún no hay calificacionesMontessori y Atención Focalizada Infantil38 páginas
- Impacto del Abuso de Drogas en la SociedadAún no hay calificacionesImpacto del Abuso de Drogas en la Sociedad69 páginas
- Corominas, M. (Et Al.) (2009) : El Sistema Dopaminérgico en Las AdiccionesAún no hay calificacionesCorominas, M. (Et Al.) (2009) : El Sistema Dopaminérgico en Las Adicciones11 páginas
- Neurociencia Afectiva y ConcienciaAún no hay calificacionesNeurociencia Afectiva y Conciencia8 páginas
- Adicción y Dopamina Rev Mente y Cerebro 2009 N 35Aún no hay calificacionesAdicción y Dopamina Rev Mente y Cerebro 2009 N 350 páginas
- Evolución y Función de la CortezaAún no hay calificacionesEvolución y Función de la Corteza19 páginas
- Psicopatología-Clínica-Basada en DSM5-Margarita-Ortiz-TalloAún no hay calificacionesPsicopatología-Clínica-Basada en DSM5-Margarita-Ortiz-Tallo8 páginas
- Factores de Riesgo en Adicción a Redes SocialesAún no hay calificacionesFactores de Riesgo en Adicción a Redes Sociales22 páginas